三、植物细胞的结构
植物细胞由原生质体(protoplast)和细胞壁(cell wall)两部分组成。原生质体是由生命物质——原生质(protoplasm)所构成,它是细胞各类代谢活动进行的主要场所,是细胞最重要的部分。细胞壁是包围在原生质体外面的坚韧外壳,长期以来,人们认为它是植物细胞的非生命部分,但近期,越来越多的研究证明,细胞壁和原生质体之间有着结构和机能上的密切联系,尤其是在幼年的细胞中,二者是一个有机的整体。
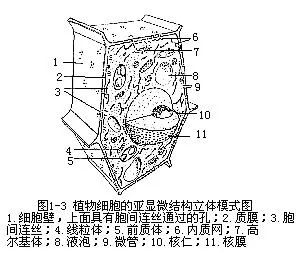
在光学显微镜下,原生质体可以明显地区分为细胞核(nucleus)和细胞质(cytopla-sm)。细胞核呈一个折光较强、粘滞性较大的球状体,与细胞质有明显的分界。细胞质是原生质体除了细胞核以外的其余部分。它们二者都不是匀质的,在内部还分化出一定的结构,其中有的用光学显微镜可以看到,而有的必须借助于电子显微镜才能显得出来。人们把在光学显微镜下呈现的细胞结构称为显微结构(microscopic structure),而将在电子显微镜下看到的更为精细的结构称为亚显微结构(submicroscopic structure)或超微结构(ultramicroscopic structure)(图1—3)。同样,细胞壁也有精细的构造。下面我们将具体地分别加以介绍。
(一)原生质体
1.细胞核 植物中除最低等的类群——细菌和蓝藻外,所有的生活细胞都具有细胞核。通常一个细胞只有一个核,但有些细胞也可以是双核或多核的,多见于菌藻植物,维管植物中少数细胞也可有二个以上的核,例如乳汁管具多核,绒毡层细胞常具二核。
细胞核的位置和形状随着细胞的生长而变化,在幼期细胞中,核位于细胞中央,近球形,并占有较大的体积。随着细胞的生长和中央液泡的形成,细胞核同细胞质一起被液泡挤向靠近壁的部位,变成半球形或圆饼状,并只占细胞总体积的一小部分。也有的细胞到成熟时,核被许多线状的细胞质索悬吊在细胞中央。然而不管是哪种情况,细胞核总是存在于细胞质中,反映出二者具有生理上的密切关系。
细胞核具有一定的结构 当观察生活细胞时,可以看到细胞核外有一层薄膜,与细胞质分界,称为核膜(nuclear membrane)。膜内充满均匀透明的胶状物质,称为核质(nucleoplasm),其中有一到几个折光强的球状小体,称为核仁(nucleolus)。当细胞固定染色后,核质中被染成深色的部分,称染色质(chromatin),其余染色浅的部分称核液(nucleochylema)。
核膜是物质进出细胞核的门户,起着控制核与细胞质之间物质交流的作用。电子显微镜观察到核膜具有双层,由外膜和内膜组成。膜上还具有许多小孔,称为核孔(nuclear pore)。这些孔能随着细胞代谢状态的不同进行启闭,所以,不仅小分子的物质能有选择地透过核膜,而且,某些大分子物质,如RNA或核糖核蛋白体颗粒等,也能通过核孔而出入,由此反映出细胞核与细胞质之间具有密切而能控制的物质交换,这种交换对调节细胞的代谢具有十分重要的作用。例如,有实验证明,小麦在活跃生长的分蘖时期,核膜上呈现相当大的孔,当进入寒冬季节时,抗寒的冬性品种的核孔,随着气温的降低逐渐关闭,而不抗寒的春性品种的核孔,却依然张开,因此,据推测,核孔的这种动态,对于小麦在低温下停止细胞分裂和生长,增进抗寒能力上起着一定的控制作用。
核仁是核内合成和贮藏RNA的场所,它的大小随细胞生理状态而变化,代谢旺盛的细胞,如分生区的细胞,往往有较大的核仁,而代谢较慢的细胞,核仁较小。
染色质是细胞中遗传物质存在的主要形式,在电子显微镜下显出一些交织成网状的细丝,主要成分是DNA和蛋白质。当细胞进行有丝分裂时,这些染色质丝便转化成粗短的染色体。
核液是核内没有明显结构的基质,化学成分尚不清楚,可能含有蛋白质、RNA和多种酶。
由于细胞内的遗传物质(DNA)主要集中在核内,因此,细胞核的主要功能是储存和传递遗传信息,在细胞遗传中起重要作用。此外,细胞核还通过控制蛋白质的合成对细胞的生理活动起着重要的调节作用,如果将核从细胞中除去,就会引起细胞代谢的不正常,并且很快导致细胞死亡。当然,细胞核生理功能的实现,也脱离不了细胞质对它的影响,细胞质中合成的物质以及来自外界的信号,也不断进入核内,使细胞核的活动作出相应的改变,因此,在细胞中,细胞核总是包埋在细胞质中的。
2.细胞质 细胞质充满在细胞核和细胞壁之间,它的外面包被着质膜(plasmalemma或plasma membrane),质膜内是透明的无结构的基质包埋着一些称为细胞器(organelle)的微小结构。细胞器是细胞质中具有一定的形态结构和具有特定功能的小“器官”,包括质体(plastid)、线粒体(mitochondria)、内质网(endoplasmic reticulum)、高尔基体(dictyosome或Golgi body)、液泡(vacuole)、微管(microtubule)……等。
(1)质膜 质膜是包围在细胞质表面的一层薄膜,在动物细胞中通常称为细胞膜(cellmembrane)。由于它很薄,通常又紧贴细胞壁,因此,在光学显微镜下较难识别。如果采用高渗溶液处理,使原生质体失水而收缩,与细胞壁发生分离(即质壁分离),就可看到质膜是一层光滑的薄膜。
在电子显微镜下,经适合的固定,质膜显出具有明显的三层结构:两侧呈两个暗带,中间夹有一个明带。三层的总厚度约7.5nm,其中两侧暗带各为2nm,中间明带约3.5nm。明带的主要成分是类脂,而暗带的主要成分为蛋白质。这种在电子显微镜下显示出由三层结构组成为一个单位的膜,称为单位膜(unit membrane),所以,质膜是一层单位膜。细胞中除质膜外,细胞核的内膜和外膜,以及其他细胞器表面的包被膜一般也都是单位膜,但各自的厚度、结构和性质都有差异。
质膜的主要功能是控制细胞与外界环境的物质交换。这是因为质膜具有“选择透性”,此种特性表现为不同的物质透过能力不同。当膜生活时,某些物质能很快透过,某些物质透过较慢,而另一些物质则不能透过。而且,随着细胞生理状态的不同,物质的这种透过能力可以发生相应的变化,在某些情况下,它们比另一些情况更能透过,这种特性是生活的生物膜所特有的。一旦细胞死亡,膜的选择透性也就随着消失,物质便能自由地透过了。质膜的选择透性使细胞能从周围环境不断地取得所需要的水分、盐类和其他必需的物质,而又阻止有害物质的进入;同时,细胞也能将代谢的废物排除出去,而又不使内部有用的成分任意流失,从而保证了细胞具有一个合适而相对稳定的内环境,这是进行正常生命活动所必需的前题。此外,质膜还有许多其他重要的生理功能,例如主动运输、接受和传递外界的信号,抵御病菌的感染,参与细胞间的相互识别等。
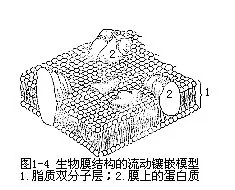
生物膜的选择透性是与它的分子结构密切相关的,但是,关于膜的分子结构,到目前为止,还没有完全被人们所了解。一般认为,磷脂是组成生物膜整体结构的主要成分,二排磷脂分子在细胞质(或细胞器)表面形成一个双分子层。在每一排中,磷脂分子与膜垂直,相互平行排列,二排分子含磷酸的亲水“头部”,分别朝向膜的内、外二侧,而疏水的脂肪酸的烃链“尾部”都朝向膜的中间,二排分子尾尾相接,这样形成了一个包围细胞质的连续脂质双分子层。生物膜就是这种脂质层与蛋白质相结合的产物。蛋白质在膜上的分布,科学家提出许多假设的模型,目前较普遍接受的二种是“膜的流动镶嵌模型”。这一学说认为,在膜上有许多球状蛋白,以各种方式镶嵌在磷脂双分子层中,有的分别结合在膜的内外表面,有的较深地嵌入磷脂质层中,再有的横向贯穿于整个双分子层(图1-4)。而且,这样的结构不是一成不变的,构成膜的磷脂和蛋白质都具有一定的流动性,可以在同一平面上自由移动,使膜的结构处于不断变动的状态。膜的选择透性主要与膜上蛋白质有关,膜蛋白大多是特异的酶类,在一定的条件下,它们具有“识别”、“捕捉”和“释放”某些物质的能力,从而对物质的透过起主要的控制作用。
(2)细胞器 细胞器一般认为是散布在细胞质内具有一定结构和功能的微结构或微器官。但对于“细胞器”这一名词的范围,还存在着某些不同意见。
①质体 质体是一类与碳水化合物的合成与贮藏密切有关的细胞器,它是植物细胞特有的结构。根据色素的不同,可将质体分成三种类型:叶绿体(chloroplast)、有色体(或称杂色体,chromoplast)和白色体(leucoplast)(图1—5)。
叶绿体是进行光合作用的质体,只存在于植物的绿色细胞中,每个细胞可以有几颗到几十颗。有人计算蓖麻的叶片每平方毫米中可有403000颗叶绿体叶。叶绿体含有叶绿素(chloro-phyll)、叶黄素(xanthophyll)和胡萝卜素(carotin),其中叶绿素是主要的光合色素,它能吸收和利用光能,直接参与光合作用。其他二类色素不能直接参与光合作用,只能将吸收的光能传递给叶绿素,起辅助光合作用的功能。植物叶片的颜色,与细胞叶绿体中这三种色素的比例有关。一般情况,叶绿素占绝对优势,叶片呈绿色,但当营养条件不良、气温降低或叶片衰老时,叶绿素含量降低,叶片便出现黄色或橙黄色。某些植物秋天叶变红色,就是因叶片细胞中的花青素和类胡萝卜素(包括叶黄素和胡萝卜素)占了优势的缘故。在农业上,常可根据叶色的变化,判断农作物的生长状况,及时采取相应的施肥、灌水等栽培措施。

高等植物的叶绿体形状相似,呈球形、卵形或凸透镜形。直径 4—10μm,厚度 1—2μm。在低等植物(藻类)中,叶绿体有各种形状,如杯状、带状和各种不规则形状。电子显微镜下显示出叶绿体具有精致的结构,表面有双层膜包被,内部有膜形成的许多圆盘状的类囊体(thy-lakoid)相互重叠,形成一个个柱状体单位,称为基粒(granum),在基粒之间,有基粒间膜(基质片层,fret)相联系。除了这些以外的其余部分是没有一定结构的基质(stroma或matrix)(图1-6A,B)。叶绿体色素位于基粒的膜上,光合作用所需的各种酶类分别定位于基粒的膜上或者在基质中,在基粒和基质中分别完成光合作用中不同的化学反应,光反应在基粒上进行,暗反应在基质中进行。

有色体只含有胡萝卜素和叶黄素,由于二者比例不同,可分别呈黄色、橙色或橙红色。它们经常存在于果实、花瓣或植物体的其他部分,例如胡萝卜的根,由于具有许多有色体而成为金黄色。有色体的形状多种多样,例如红辣椒果皮中的有色体呈颗粒状,旱金莲花瓣中的有色体呈针状。有色体能积聚淀粉和脂类,在花和果实中具有吸引昆虫和其他动物传粉及传播种子的作用。
白色体不含色素,呈无色颗粒状。普遍存在于植物体各部分的储藏细胞中,起着淀粉和脂肪合成中心的作用。当白色体特化成淀粉储藏体时,便称为淀粉体(amyloplast),当它形成脂肪时,则称为造油体(elaioplast)。
在电子显微镜下,可以看到有色体和白色体表面也有双层膜包被,但内部没有发达的膜结构,不形成基粒。
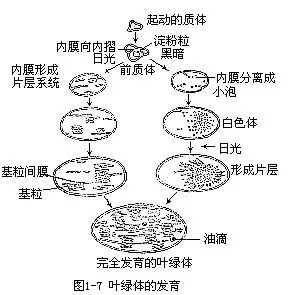
关于质体的发育,一般认为是由幼小细胞中的前质体(proplastid)发育而来的。前质体是一种较小的无色体,能分裂。直径约 1—3μm。最初,在幼小细胞内有一些为双层膜所包被的小泡,其中没有片层结构,以后,小泡内膜向内折叠,内折的膜层与小泡表面平行,这时称为前质体。前质体进一步的发育,因外界条件而异,在光照条件下,内膜逐渐发育成正常的叶绿体基粒,同时形成叶绿素,发育为叶绿体;而在黑暗的条件下,内膜形成分离的管子,相互连接成立体的网格,同时也不形成色素,发育成白色体。这是黑暗中生长的植物会出现黄化的原因。如果黄化的植物再转入光下,白色体又可以发育成正常的叶绿体(图1-7)。有色体一般认为不是由前质体直接发育而来的,它是由白色体或叶绿体转化而成的。例如发育中的番茄,最初含有白色体,以后转化成叶绿体,最后,叶绿体失去叶绿素而转化成有色体,果实的颜色也随着变化,从白色变成绿色,最后成为红色。相反,有色体也能转化成其他质体,例如,胡萝卜根的有色体暴露于光下,就可发育为叶绿体。
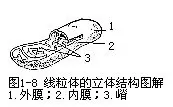
②线粒体 线粒体是一些大小不一的球状、棒状或细丝状颗粒,一般直径为0.5—1μm,长度是1-2μm,在光学显微镜下,需用特殊的染色,才能加以辨别。在电子显微镜下可看出,线粒体由双层膜包裹着,其内膜向中心腔内折叠,形成许多隔板状或管状突起,称为嵴(cris-tae)。在二层被膜之间及中心腔内,是以可溶性蛋白为主的基质(图1-8)。
线粒体是细胞进行呼吸作用的场所,它具有100多种酶,分别存在于膜上和基质中,其中极大部分参与呼吸作用。线粒体呼吸释放的能量,能透过膜转运到细胞的其他部分,提供各种代谢活动的需要,因此,线粒体被比喻为细胞中的“动力工厂”。
细胞中线粒体的数目,以及线粒体中嵴的多少,与细胞的生理状态有关。当代谢旺盛,能量消耗多时,细胞就具有较多的线粒体,其内有较密的嵴;反之,代谢较弱的细胞,线粒体较少,内部嵴也较疏。
③内质网 内质网是分布于细胞质中由一层膜构成的网状管道系统,管道以各种形状延伸和扩展,成为各类管、泡、腔交织的状态。在超薄切片中,内质网看起来是二层平行的膜,中间夹有一个窄的空间。每层膜的厚度约为5nm,二层膜之间的距离只有40—70nm,必须借助电子显微镜才能辨别(图1-9,A;图1-10)。
内质网有两种类型,一类在膜的外侧附有许多小颗粒,这种附有颗粒的内质网称为粗糙型内质网(图1—9,A),这些颗粒是核糖核蛋白体(ribosome);另一类在膜的外侧不附有颗粒,表面光滑,称光滑型内质网。细胞中,二类内质网的比例及它们的总量,随着细胞的发育时期、细胞的功能和外部条件而变化。核糖核蛋白体也称核蛋白体或核糖体。
关于内质网的功能,目前还未完全清楚。在细胞中,内质网可以与细胞核的外膜相连,同时,也可与原生质体表面的质膜相连,有的还随同胞间连丝穿过细胞壁,与相邻细胞的内质网发生联系。因此,有人认为内质网构成了一个从细胞核到质膜,以及与相邻细胞直接相通的管道系统,与细胞内和细胞间的物质运输有关。粗糙型内质网与核糖核蛋白体紧密结合,而核糖核蛋白体是合成蛋白质的细胞器,因此,推测粗糙型内质网与蛋白质(主要是酶)合成有关,膜上核糖核蛋白体合成的蛋白质进入内质网腔内,并进一步将它转运到细胞其他部位去。光滑型内质网主要合成和运输类脂和多糖,例如,在分泌脂类物质的细胞中,常常有广泛的光滑型内质网。在细胞壁进行次生增厚的部位内方,也可以见到内质网紧靠质膜,反映了内质网可能与加到壁上去的多糖类的合成有关。

④高尔基体 高尔基体是由一叠扁平的囊(cisterna,也称为泡囊或槽库)所组成的结构,每个囊由单层膜包围而成,直径约0.5—1μm,中央似盘底,边缘或多或少出现穿孔。当穿孔扩大时,囊的边缘便显得象网状的结构。在网状部分的外侧,局部区域膨大,形成小泡(vesi-cle),通过缢缩断裂,小泡从高尔基体囊上可分离出去(图1-9,B;图1-10)。

高尔基体与细胞的分泌功能相联系。分泌物可以在高尔基体中合成,或来源于其他部分(如内质网),经高尔基体进一步加工后,再由高尔基小泡将它们携带转运到目的地。分泌物主要是多糖和多糖-蛋白质复合体。这些物质主要用来提供细胞壁的生长,或分泌到细胞外面去。当小泡输送物质参与壁的生长时,小泡向质膜移动,先与质膜接触,二者的膜发生融合,然后小泡内容物向壁释放出去,添加到壁上。在有丝分裂形成新细胞壁的过程中,可以看到大量高尔基小泡,运送形成新壁所需要的多糖类物质,参与新细胞壁的形成。也有实验证明,根的根冠(root cap)细胞分泌粘液,松树的树脂道(resin canal)上皮细胞分泌树脂等,也都与高尔基体活动有关。一个细胞内的全部高尔基体,总称为高尔基器(Golgi appaatus)。
⑤核糖核蛋白体 核糖核蛋白体简称为核糖体,是直径为17—23nm的小椭圆形颗粒(图1-10)。它的主要成分是RNA和蛋白质。在细胞质中,它们可以游离状态存在,也可以附着于粗糙型内质网的膜上。此外,在细胞核、线粒体和叶绿体中也存在。
核糖核蛋白体是细胞中蛋白质合成的中心,氨基酸在它上面有规则地组装成蛋白质。所以,蛋白质合成旺盛的细胞,尤其在快速增殖的细胞中,往往含有更多的核糖核蛋白体颗粒。
在执行蛋白质合成功能时,单个核糖核蛋白体经常5—6个或更多个串联在一起,形成一个聚合体,称为多核蛋白体或多核糖体(polyribosome或polysome),它的合成效率比单个的更高。
⑥液泡 具有一个大的中央液泡是成熟的植物生活细胞的显著特征,也是植物细胞与动物细胞在结构上的明显区别之一。
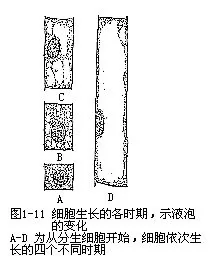
幼小的植物细胞(分生组织细胞),具有许多小而分散的液泡,它们在电子显微镜下才能看到。以后,随着细胞的生长,液泡也长大,相互并合,最后在细胞中央形成一个大的中央液泡,它可占据细胞体积的90%以上。这时,细胞质的其余部分,连同细胞核一起,被挤成为紧贴细胞壁的一个薄层(图l-11)。有些细胞成熟时,也可以同时保留几个较大的液泡,这样,细胞核就被液泡所分割成的细胞质索悬挂于细胞的中央。
液泡是被一层液泡膜(tonoplast)包被,膜内充满着细胞液(cell sap),它是含有多种有机物和无机物的复杂的水溶液。这些物质中有的是细胞代谢产生的储藏物,例如糖、有机酸、蛋白质、磷脂等;有的是排泄物,例如草酸钙、花色素(anthocyanidin)等,但是,在植物中储藏物和排泄物没有严格的界线,因为植物具有对物质再度转化利用的能力。液泡膜具有特殊的选择透性,能使许多物质大量积聚在液泡中。例如甘蔗的茎和甜菜的根中,含有大量蔗糖,具有浓厚的甜味。许多果实含有丰富的有机酸,造成强烈的酸味。茶叶和柿子等因含大量丹宁而具涩味,并使破损后伤口很快变成黑色。许多植物含丰富的植物碱,如罂粟(Papaversomniferum)含吗啡,烟草含尼古丁,茶叶和咖啡含咖啡碱等。许多植物细胞液中溶解有花色素,从而使花瓣、果实或叶片显出红色、紫色或蓝色。花色素的显色与细胞液pH有关,酸性时呈红色,碱性时呈蓝色,中性时呈紫色,常见的牵牛花在早晨为蓝色,以后渐转红色,就是这个缘故。细胞液还含有很多无机盐,有些盐类因过饱和而成结晶,常见的如草酸钙结晶。细胞液中各类物质的富集,使细胞液保持相当的浓度,这对于细胞渗透压和膨压的维持,以及水分的吸收有着很大的关系,使细胞能保持一定的形状和进行正常的活动。同时,高浓度的细胞液,使细胞在低温时不易冻结,在干旱时不易丧失水分,提高了抗寒和抗旱的能力。
电子显微镜的观察和酶的定位研究,进一步证明液泡不仅能单纯地储藏代谢产物,而且也积极地参与细胞中物质的生化循环,参与细胞分化和细胞衰老等重要的生命过程。液泡中具有不少酶类,其中包括多种水解酶,它们在一定条件下,能分解液泡中的储藏物质,重新动用来参加各种代谢活动。在电子显微镜下,经常还可以看到薄壁细胞的正常液泡中,悬浮有不完整的线粒体、质体或内质网片段等,它们是被液泡膜“吞噬”进去的细胞衰老的组成部分,以后在液泡中被分解而消失,这是细胞结构新陈代谢的一种方式,是细胞分化及衰老、死亡过程中必需的过程。因此,液泡被越来越多的人认为也是一种具有重要生理功能的细胞器,是细胞质的一部分。
植物细胞具有大的中央液泡,可能有着一种特殊的生理意义。细胞代谢所需要的物质,必须通过细胞表面进入细胞,但植物细胞具有坚固的细胞壁,它不能像动物细胞那样,通过改变细胞的形状来扩大吸收表面,因此,它通过另外的方式来达到相似的目的,即借助于大的中央液泡,把细胞质挤压成贴壁的薄层,这样,便有利于原生质体与外界发生气体和养料的交换。
液泡中的代谢产物不仅对植物细胞本身具有重要的生理意义,而且,植物液泡中丰富而多样的代谢产物是人们开发利用植物资源的重要来源之一,例如,从甘蔗的茎、甜菜的根中提取蔗糖,从罂粟果实中提取鸦片,从盐肤木、化香树中提取单宁作为烤胶的原料等。近年来,开发新的野生植物资源正在引起人们越来越大的兴趣,如刺梨、酸枣等果实被用作制取新型饮料;从花、果实中提取天然色素,用于轻工、化工,尤其是食品工业的着色。天然色素的开发已成为当前国内外十分重视的一个研究领域。
⑦溶酶体(lysosome)溶酶体是由单层膜包围的多形小泡,一般直径为 0.25—0.3μm。内部主要含有各种不同的水解酶类,如酸性磷酸酶、核糖核酸酶、组织蛋白酶、脂酶等,它们能分解所有的生物大分子,“溶酶体”因此而得名,溶酶体可以通过膜的内陷,把细胞质的其他组分吞噬进去,在溶酶体内进行消化;也可通过本身膜的解体,把酶释放到细胞质中而起作用。溶酶体在细胞内对贮藏物质的利用起重要作用,同时,在细胞分化过程中对消除不必要的结构组成,以及在细胞衰老过程中破坏原生质体结构也都起特定的作用。例如,在导管和纤维成熟时,原生质体最后完全破坏消失,这一过程就与溶酶体的作用密切有关。
“溶酶体”最早的概念来自动物细胞,指含有水解酶类的细胞器。现在了解,在植物细胞中,许多结构组分都含有酸性水解酶类,都具有分解代谢物及细胞组分的本领,其中如液泡就占据重要的地位。另外,糊粉粒、圆球体等也都含水解酶,具有溶酶体的作用。因此,一些人认为,在植物细胞中溶酶体不是一个特殊的形态学实体,而应指能发生水解作用的所有结构。
⑧圆球体(spherosome)圆球体是膜包裹着的圆球状小体,直径为0.1—1μm,染色反应似脂肪,用锇酸固定后成为或多或少深色的球体。电子显微镜观察指出,它的膜只具有一层电子不透明层(暗带),而不象正常的单位膜具二个暗带,因此,可能只是单位膜的一半。膜内部有一些细微的颗粒结构。圆球体是一种储藏细胞器,是腊肪积累的场所,当大量脂肪积累后,圆球体便变成透明的油滴,内部颗粒消失。在圆球体中也检定出含有脂肪酶,在一定条件下,酶也能将脂肪水解成甘油和脂肪酸。因此,圆球体具有溶酶体的性质。
⑨微体(microbody或 cytosome)微体是一些由单层膜包围的小体,直径约0.5μm。它的大小、形状与溶酶体相似,二者的区别在于含有不同的酶。微体含有氧化酶和过氧化氢酶类。另外,有些微体中含有小的颗粒、纤丝或晶体等(图1-12)。

现在了解到微体有二种:过氧化物酶体(peroxisome)和乙醛酸循环体(glyoxysome)。过氧化物酶体存在于高等植物叶肉细胞内,它与叶绿体、线粒体相配合,参与乙醇酸循环,将光合作用过程中产生的乙醇酸转化成已糖。乙醛酸循环体主要出现在油料种子萌发时,它与圆球体和线粒体相配合,把储藏的脂肪转化成糖类。
⑩微管和微丝(microfilament)微管和微丝是细胞内呈管状或纤丝状的二类细胞器,它们在细胞中相互交织,形成一个网状的结构,成为细胞内的骨骼状的支架,使细胞具有一定的形状,在细胞学上称它们为微梁系统(microtrabecularsystem)。

微管在电子显微镜下是中空而直的细管,长约数微米,直径约25nm,其中管壁厚4—5nm,中心是电子透明的空腔(图1-13)。微管的化学组成是微管蛋白,它是一种球蛋白,在细胞中能随着不同的条件迅速地装配成微管,或又很快地解聚,因而,微管成为一种不稳定的细胞器。在低温、压力、秋水仙碱(colchicine)、酶等外界条件的作用下,微管也很容易被破坏。
微管的生理功能主要有几个方面:第一,微管与细胞形状的维持有一定的关系。有人研究植物呈纺锤状的精子,发现它具有与细胞长轴相平行的微管,当用秋水仙碱处理后,微管被破坏,精子便变成球形。因此,说明微管可能在细胞中起支架作用,保持细胞一定的形状;第二,微管参与细胞壁的形成和生长。在细胞分裂时,由微管组成的成膜体,指导着含有多糖类物质的高尔基体小泡,向新细胞壁方向运动,在赤道面集中,融合形成细胞板。其后,微管在质膜下的排列方向,又决定着细胞壁上纤维素微纤丝的沉积方向。并且,在细胞壁进一步增厚时,微管集中的部位与细胞壁增厚的部位是相应的,反映了壁的增厚方式可能也受微管的控制;第三,微管与细胞的运动及细胞内细胞器的运动有密切关系。植物游动细胞的纤毛和鞭毛,是由微管构成的。在细胞分裂时,染色体的运动受微管构成的纺锤丝的控制。也有实验指出,细胞内细胞器的运动方向,也受微管的控制。
微丝是比微管更细的纤丝,直径只有5—8nm。在细胞中呈纵横交织的网状,与微管共同构成细胞内的支架,维持细胞的形状,并支持和网络各类细胞器。例如“游离”的核糖核蛋白体颗粒,很可能是连接在此网络的交叉点上。
微丝的主要成分是类似于肌动蛋白和肌球蛋白的蛋白质,因此,它具有像肌肉一样的收缩功能,除了起支架作用外,它的主要功能是与微管配合,控制细胞器的运动,微管的排列为细胞器提供了运动的方向,而微丝的收缩功能,直接导致了运动的实现。另外,微丝与胞质流动(cytoplasmic streaming)有密切的关系。在具有明显的胞质流动的细胞中,可以看到成束的微丝排列在流动带中,并与流动方向相平行,当用专门破坏微丝的药品——细胞松弛素处理后,胞质流动便停止。如果把药物去掉,微丝可重新聚合,胞质流动又可恢复。
(3)胞基质(cytoplasmic matrix)在电子显微镜下,看不出特殊结构的细胞质部分,称为胞基质。细胞器及细胞核都包埋于其中。它的化学成分很复杂,包含水、无机盐、溶解的气体、糖类、氨基酸,核苷酸等小分子物质,也含有一些生物大分子,如蛋白质、RNA等,其中包括许多酶类。它们使胞基质表现为具有一定弹性和粘滞性的胶体溶液,而且它的粘滞性可随着细胞生理状态的不同而发生改变。
在生活的细胞中,胞基质处于不断的运动状态,它能带动其中的细胞器,在细胞内作有规则的持续的流动,这种运动称胞质运动(cytoplasmic movement)。在具有单个大液泡的细胞中,胞基质常常围绕着液泡朝一个方向作循环流动。而在具有多个液泡的细胞中,不同的细胞质索可以有不同的流动方向。胞质运动是一种消耗能量的生命现象,它的速度与细胞生理状态有密切的关系,一旦细胞死亡,流动也随着停止。胞质运动对于细胞内物质的转运具有重要的作用,促进了细胞器之间生理上的相互联系。
胞基质不仅是细胞器之间物质运输和信息传递的介质,而且也是细胞代谢的一个重要场所,许多生化反应,如厌氧呼吸及某些蛋白质的合成等就是在胞基质中进行的。同时,胞基质也不断为各类细胞器行使功能提供必需的原料。
综上所述,我们可以看到植物细胞的原生质体,是细胞内一团结构上具有复杂分化的原生质单位。在细胞质膜内,具有特定形态和功能的细胞器,悬浮于以蛋白质为主的胶状基质中。从细胞器的定义出发,细胞核也可作为一个控制细胞遗传和发育的特殊的细胞器。这些结构,在功能上具有分工,但又是相互联系,相互依赖的,例如植物细胞最基本的生命活动——呼吸作用是在线粒体中进行的,然而呼吸所需的物质基础必须依赖叶绿体的光合作用提供;参与呼吸作用的各种酶类必须由核糖体合成;呼吸产生的能量必须通过胞基质转运到其他细胞器中,供各类代谢活动的需要,而水和二氧化碳又必须借助于质膜排出体外,或重新用作光合作用的原料。同时,参与以上种种代谢活动的酶的合成,又必然要受到细胞核的控制。由此可见,原生质体总是作为一个整体单位而进行生命活动的。
各类细胞器不仅功能上密切联系,而且在结构上和起源上也是相联系的。绝大部分的细胞器都是由膜所围成,各类细胞器的膜在成分上和功能上虽具有各自的特异性,但它们的基本结构是相似的,都是单位膜。它们在细胞内发育上的联系,可以用图1-14来作个示意:核膜的外膜与粗糙型内质网相联系,光滑型内质网产生的囊泡可以转化为高尔基体的泡囊,内质网和高尔基体又可以发育出液泡和各类小泡,小泡又可进一步发育为溶酶体、圆球体和微体等。因此,许多生物学家认为,细胞内这些细胞器是一个统一的、相互联系的膜系统在局部区域特化的结果,这个膜系统称为细胞的内膜系统。“内膜”是相对于包围在外面的质膜而言的。
在生物进化过程中,内膜系统在原生质体中起分隔化、区域化的作用。被膜分隔的不同小区,特化为不同的细胞器,从而实现细胞内的区域分工,使得在“细胞”——这样一个极小的空间中能同时进行多种不同的生化反应。内膜系统巨大的表面,又使各种酶能定位于不同的部位,保证了一系列复杂的生化反应能有顺序地、高效地进行。同时,内膜系统还与质膜相连,相邻细胞的内膜系统通过胞间联丝也互相沟通,这就提供了一个细胞内及细胞间的物质和信息的运输系统,从而使多细胞有机体能成为协调的统一整体。
因此,具有内膜系统是生物进化的表现,只有真核细胞(即具有真正细胞核的细胞)才有内膜系统,处于较低级状态的原核细胞(即没有真正细胞核的细胞)不分化成内膜系统。


(二)细胞壁
细胞壁是包围在植物细胞原生质体外面的一个坚韧的外壳。它是植物细胞特有的结构,与液泡、质体一起构成了植物细胞与动物细胞相区别的三大结构特征。
细胞壁的功能是对原生质体起保护作用。此外,在多细胞植物体中,各类不同的细胞的壁,具有不同的厚度和成分,从而影响着植物的吸收、保护、支持、蒸腾和物质运输等重要的生理活动。有人将细胞壁比喻成植物的皮肤、骨骼和循环系统。
一般认为,细胞壁在本质上不是一种生活系统,它是由原生质体分泌的非生活物质所构成的,但是细胞壁与原生质体又保持有密切的联系。在年幼的细胞中,细胞壁与原生质体紧密结合,即使用较高浓度的糖溶液,也不能引起质壁分离。现在已经证明,在细胞壁(主要是初生壁)中亦含有多种具有生理活性的蛋白质,它们可能参与细胞壁的生长、物质的吸收、细胞间的相互识别以及细胞分化时壁的分解等过程,有的还对抵御病原菌的入侵起重要作用。
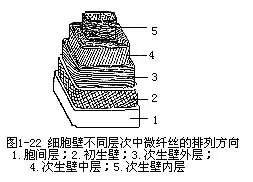
1.细胞壁的层次 细胞壁根据形成的时间和化学成分的不同分成三层:胞间层(intercel-lular layer)、初生壁(Primary wall)和次生壁(secondary wall)(图1-15)。
(1)胞间层 又称中层,存在于细胞壁的最外面。它的化学成分主要是果胶(pectin),这是一种无定形胶质,有很强的亲水性和可塑性,多细胞植物依靠它使相邻细胞彼此粘连在一起。果胶很易被酸或酶等溶解,从而导致细胞的相互分离。例如某些组织成熟时,体内的酶分解部分胞间层,形成细胞间隙。许多果实,如番茄、苹果、西瓜等成熟时,果肉细胞的胞间层被溶解,致使细胞发生分离,果肉变得软而“面”。有些真菌能分泌果胶酶,溶解植物组织的胞间层而侵入植物体内。麻类植物的茎浸入水中的沤麻过程,也是利用微生物分泌酶分解纤维的胞间层使其相互分离。
(2)初生壁 初生壁是在细胞停止生长前原生质体分泌形成的细胞壁层,存在于胞间层内侧。它的主要成分是纤维素、半纤维素和果胶。现已证明,在初生壁中也含有少量结构蛋白,这些蛋白质成分与壁上的多糖紧密结合。初生壁的厚度一般较薄,约1—3μm,质地较柔软,有较大的可塑性,能随着细胞的生长而延展。许多细胞在形成初生壁后,如不再有新壁层的积累,初生壁便成为它们永久的细胞壁。
(3)次生壁 次生壁是细胞停止生长后,在初生壁内侧继续积累的细胞壁层。它的主要成分是纤维素(cellulose),含有少量的半纤维素(hemicellulose),并常常含有木质(lignin)。次生壁较厚,一般约5—10μm,质地较坚硬,因此,有增强细胞壁机械强度的作用。在光学显微镜下,厚的次生壁层可以显出折光不同的三层:外层、中层和内层。因此,一个典型的具次生壁的厚壁细胞(如纤维或石细胞),细胞壁可看到有5层结构:胞间层、初生壁和三层次生壁。但是,不是所有的细胞都具有次生壁,大部分具次生壁的细胞,在成熟时原生质体死亡,残留的细胞壁起支持和保护植物体的功能。
2.纹孔(pit)和胞间连丝(plasmodesmata)细胞壁生长时并不是均匀增厚的。在初生壁上具有一些明显的凹陷区域,称为初生纹孔场(primary pit field)。在初生纹孔场上集中分布着许多小孔,细胞的原生质细丝通过这些小孔,与相邻细胞的原生质体相连。这种穿过细胞壁,沟通相邻细胞的原生质细丝称为胞间连丝(图1-16;图1-17,A),它是细胞原生质体之间物质和信息直接联系的桥梁,是多细胞植物体成为一个结构和功能上统一的有机体的重要保证。在高倍电子显微镜下,胞间连丝显出是直径约40nm的管状结构,相邻细胞的质膜在胞间连丝周围是连续的,丝的中心可观察到直径为10nm的深色的结构,内质网通过胞间连丝与相邻细胞联系。除初生纹孔场外,在壁的其他部位也可分散存在少量胞间连丝。


当次生壁形成时,次生壁上具有一些中断的部分,这些部分也就是初生壁完全不被次生壁覆盖的区域,称为纹孔。纹孔如在初生纹孔场上形成,一个初生纹孔场上可有几个纹孔。一个纹孔由纹孔腔(pit cavity)和纹孔膜(pit membrane)组成,纹孔腔是指次生壁围成的腔,它的开口(纹孔口)朝向细胞腔。腔底的初生壁和胞间层部分即称纹孔膜。根据次生壁增厚情况的不同,纹孔分成单纹孔(simple pit)和具缘纹孔(bordered pit)两种类型,它们的基本区别是具缘纹孔的次生壁穹出于纹孔腔上,形成一个穹形的边缘,从而使纹孔口明显变小,而单纹孔的次生壁没有这样的穹形边缘(图1-17,B-G),在某些裸子植物管胞的壁上有一种较为特殊的具缘纹孔,它们的纹孔膜中央部位有一个圆盘状的增厚区域,称纹孔塞(torus)。它的直径大于纹孔口,纹孔塞的增厚是初生壁性质的,周围的纹孔膜具孔称塞缘(或塞周膜,margo)。其多孔结构,有较好的透性。当纹孔膜位于相邻细胞的中央位置时,水分主要通过塞缘透到相邻细胞,而当两侧的细胞内压力不同时,纹孔膜偏向压力小的一侧,从而使纹孔塞关闭了该侧的纹孔口,阻止了水流向该侧的流动(图1-18)。可见这种具缘纹孔在一定条件下有控制水流的作用。

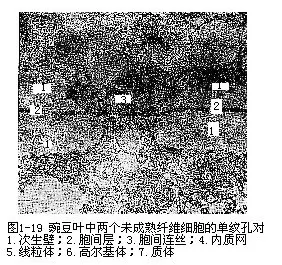
细胞壁上的纹孔通常与相邻细胞壁上的一个纹孔相对,二个相对的纹孔合称纹孔对(pitpair),纹孔对中的纹孔膜是由二层初生壁和一层胞间层组成(图1-19)。
细胞壁上初生纹孔场、纹孔和胞间连丝的存在,都有利于细胞与环境以及细胞之间的物质交流,尤其是胞间连丝,它把所有生活细胞的原生质体连接成一个整体,从而使多细胞植物在结构和生理活动上成为一个统一的有机体。
3.细胞壁的化学组成 细胞壁最主要的化学成分为纤维素,它是一种亲水的具有某些晶体性质的化合物,由100个或更多个葡萄糖基连接而成,分子成长短不等的链状。与纤维素相结合普遍存在于壁中的其他化合物是果胶质、半纤维素和非纤维素多糖。由于这些物质都是亲水性的,因此,细胞壁中一般含有较多的水分,溶于水中的任何物质都能随水透过细胞壁。
但是,在植物体中,不同细胞的细胞壁组分变化很大,这是由于细胞壁中还渗入了其他各种物质的结果。常见的物质有角质、栓质、木质、矿质等,它们渗入细胞壁的过程分别称为角质化、栓质比、木质化和矿质化。由于这些物质的性质不同,从而使各种细胞壁具有不同的性质,例如,角质和栓质是脂肪性物质,因此,角质化或栓质化的壁就不易透水,具有减少蒸腾和免于雨水浸渍的作用;木质是亲水性的,它有很大的硬度,因此,木质化的壁既加强了机械强度,又能透水;矿质主要是碳酸钙和硅化物,矿质化的壁也具有大的硬度,增加了支持力。细胞壁成分和性质上的这些差异,对于不同细胞更好地适应它所执行的功能,具有重要意义。例如茎、叶表面的细胞,细胞壁常角质化或栓质化,使植物在烈日下减少体内水分的丧失,加强了保护作用;树木木质部的细胞强烈木质化,使茎干能承受大的压力,加强了支持功能等等。

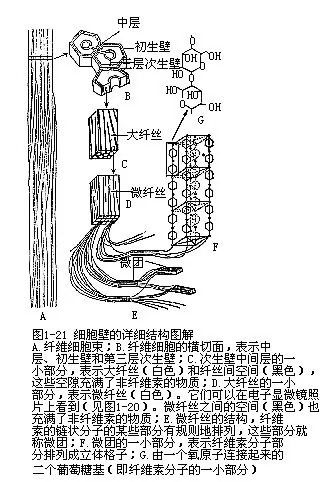
4.细胞壁的亚显微结构 电子显微镜下对细胞壁结构的研究指出,构成细胞壁的结构单位是微纤丝(microfibril)。微纤丝是由纤维素分子束(微团,micelle)聚合成的纤丝,在电子显微镜下可以辨别。把细胞壁中的非纤维素成分去掉后,可以看到微纤丝相互交织成网状,构成了细胞壁的基本构架(图1—20)。在完整的壁中,其他的壁物质(果胶、半纤维素、木质、栓质等)填充于微纤丝“网”的空隙中。微纤丝再聚集成较粗的纤丝而称为大纤丝(macrofibril),这种大纤丝可以在光学显微镜下看到(图1-21)。
在细胞壁生长时,微纤丝是成层连续地敷设到壁上去的,微纤丝的沉积方向很可能受到质膜下微管排列方向的控制,因为它们二者常常反映出具有一致性。而微纤丝的合成,则可能与质膜有关。
成熟细胞中,不同细胞壁层的微纤丝,排列成不同的方向,在初生壁中,微纤丝呈网状排列,但多数纤丝与细胞的长轴相平行;在次生壁中,微纤丝多数与长轴成一定角度斜向排列,而且,在次生壁的外、中、内三层中,微纤丝的走向也不一致,这样的排列方式大大增强了细胞壁的坚固性(图1-22)。在显微镜下,我们看到厚壁细胞和管胞的次生壁显出三层也就是这个原因。