一、植物的起源
植物界不仅类群繁多,形态多种多样,而且分布也极广泛。然而,当地球形成之初,表面非常炽热,而且外面还没有大气圈,不具备产生生命物质的条件,直到以后,地球表面出现了大气层,生命的出现才有了可能。
估计在40—35亿年前就开始出现了原始的生物,最先出现的植物是细菌和蓝藻,这两类植物的形态结构都非常简单,它们没有定型的细胞核,也没有质体和其他细胞器。因此,它们被称为原核生物(Procaryote)。蓝藻和一部分具有色素的细菌在一起,利用日光以制造养料,并放出大量的氧气,从而逐渐改变大气的性质,使它由还原性变为氧化性,这样,就为喜氧植物的出现准备了条件。
到距今15—14亿年前,开始出现了具有真核细胞的藻类。它们有定型的细胞核和细胞器,这一转变在植物界的进化途中是一次巨大的飞跃。由于细胞器的出现,使细胞内各部分的分工更为明确,从而提高了整个细胞生理活动的机能。
最初出现的真核生物,可能是生活在水中的鞭毛生物。鞭毛生物是单细胞体,它们有一根或两根鞭毛,可以自由活动,有的细胞,体外具有1层膜,能改变体形。在以后进一步的系统发育过程中,鞭毛生物体表产生1层坚固的厚壁,此厚壁是由与纤维素相近似的物质所组成;有的鞭毛生物具有典型的色素体,这与细菌和蓝藻有着很大的不同。鞭毛生物有具色素体能独立营养的植物类型,也有藉吞食现成有机物营养的动物类型。从能独立营养的鞭毛生物,又不断演化成多种多样的藻类植物。
到距今9—7亿年前,便开始出现了多细胞的藻类。最初的多细胞藻是丝状体的类型,到6亿年前又出现了囊状、柱状或其他形状的类型,这些藻类分别属于绿藻门、红藻门、褐藻门,它们不但在体形上和大小上有千差万别的变化,就是在内部结构上也日趋复杂化。这些藻类植物群主要生活在海洋里,因此,从太古代到古生代的志留纪中期,为海产藻类繁盛时期。由于藻类植物群在海洋中的大量繁衍,他们在光合作用的过程中,放出了大量的氧气,这不仅使大气的成分逐渐改变,也使海水中的含氧量增高,有利于海洋动、植物的生存。另外,也由于一部分的氧在大气上层形成了臭氧(O3),便阻挡了杀伤力甚强的紫外线辐射,从而使植物从海水登上陆地生活成为可能。在志留纪晚期,一批生于水中的裸蕨类植物,开始逐渐地进入了陆地,这是植物进化史中的一次重大的飞跃。登陆了的裸蕨类植物,进一步向适应陆生生活方向演进。到古生代的石炭纪,世界各地出现了参天的茂密蕨类植物森林,而这些早期出现的种类繁多的裸蕨类植物,却在泥盆纪末期、石炭纪以前消逝了。从石炭纪到二叠纪下期这一段地质年代中,是蕨类植物鼎盛时期,此期中有的蕨类遗体大量地被埋到地下,年长日久形成煤层,被现代人挖掘出来作为能源利用。在蕨类植物繁盛时期的同时,苔藓植物也以其独特的生活方式。成功地适应着陆生生活,繁茂地生长着。在古生代末期的二叠纪时,由于地球上出现了明显的气候带,许多地区变得不适于蕨类植物的生长,多数蕨类植物开始走向衰亡。裸子植物开始兴起,逐渐地取代了蕨类植物的优势。由古生代末期的二叠纪到中生代的白垩纪早期,这长达1亿年之久的历史期间,是裸子植物繁盛时期。裸子植物取代了蕨类植物为地球上优势植物类群,高大的裸子植物广布南北半球的各个气候带。到中生代末期,距今约1亿年前后,地球上气候带分带现象更趋明显,而且后来又出现了几次冰川时期,气温大幅度地下降,在这严酷的环境下,裸子植物类群中,多数种类由于不能适应气候的变化,逐渐消逝了,代之而起的是被子植物。被子植物与裸子植物相比,有更优越的适应环境能力,主要表现在其繁殖器官的结构与机能方面;另外,也表现在其营养体生活的多样化,如木本、草本、一年生、多年生等多方面。因此,在此时期,被子植物成了世界上的优势植物群,而代替了以前的裸子植物群,直至今日(表6-1)。
表6-1 地质年代和不同时期占优势的植物和进化情况

二、植物营养体的演化
孢子植物营养体的形态和结构是多种多样的,但它们遵循着演化规律,由简单到复杂,由低级到高级地向前发展着。一般认为,单细胞鞭毛藻类是植物界中最简单、最原始的类型,也是藻类植物和一切高等植物的祖先。
植物营养体的演化过程,在绿藻植物中最为明显,各种类型都具备(参看藻类小结)。像衣藻、盘藻、实球藻、空球藻、杂球藻到团藻,是代表着能活动的、营养体细胞有定数的、营养时期不分裂的类型。由单细胞至群体再到多细胞的发展过程中,到团藻成为这一路线的顶端,然而这一路线在植物界中是不可能得到发展的。另外,像绿球藻目植物,营养体不能活动,细胞有定数的类型,其中有些种类(如水网藻)的营养体细胞也能分裂,成为多核细胞的群体,但这条路线同样也是不可能发展的。另一类营养体为多细胞,失去活动能力,而成丝状体的类型(如丝藻目植物)成为植物界发展的主干,进一步由丝状体到异丝状体和片状体,并由此发展出高等植物来。
在褐藻门和红藻门中,较高级的种类,其体型更为复杂,有的分化成类似高等植物的根、茎、叶的体型,并有类似组织的结构。像褐藻门中的马尾藻属、巨藻属、海棕榈属(Postelsia)和红藻门中的红叶藻属(Delesseria)等。但褐藻门和红藻门植物营养体的发展进程,仍然是和绿藻门相似,遵循着从单细胞到多细胞、能游动到不能游动、分化简单到分化复杂的路线发展,这是一种和绿藻平行发展的结果。
高等植物的营养体都是多细胞的,苔类植物一般为叶状体,有背腹之分,具有单细胞的假根,而苔类以上植物已具有了气孔的结构,合子萌发也必须经历胚的阶段。藓类具有“叶”和“茎”的辐射对称的拟茎叶体,假根为多细胞。到了蕨类植物,营养体有了更进一步的演化,具有真正的根、茎、叶等器官和较完善的组织构造,特别是具有了适应陆生生活的输导系统,中柱类型复杂,由原生中柱向网状中柱发展,茎、叶上有毛或鳞片等附属物保护。叶脉由叉状分枝发展到网状分枝,孢子囊由枝端向叶缘到叶背着生等等各种类型,使植物体更能向适应环境而演变。到了种子植物,营养体变得更为多样化,内部结构也更趋完善。
三、有性生殖方式的进化
一切生物都有繁殖后代的能力,原始的细菌、蓝藻植物是以营养繁殖和无性生殖来繁衍后代的,而真核植物则普遍存在着有性生殖的繁殖方式。有性生殖有同配、异配和卵式配合3种类型。
在同配生殖中,雌、雄配子的形态、大小几乎一样而难以区分。这种生殖类型又可分为同宗配合和异宗配合两类,如某些真菌的菌丝体是行同宗配合的,而衣藻属有同宗配合的,也有异宗配合的,到盘藻、水网藻、丝藻等则均属异宗配合的类型。异宗配合比同宗配合在细胞分化上是较进化的。
异配生殖的两种配子在形态、大小上均有区别,像空球藻的有性生殖在产生雄配子时,每个细胞经纵分裂成64个细长的配子,每个配子有2根鞭毛,能单独游动;产生雌配子的是由1个不经分裂的普通细胞转变而成,而且比雄配子大好多倍,形状也不同,也不能脱离母体行单独游动。
卵式生殖是精子和卵的受精过程,在植物界中是一种最进化的有性生殖形式。
从有性生殖进化的过程来看,同配生殖是最为原始的,异配生殖其次,卵式生殖最为高等。这从团藻目植物如盘藻、实球藻、空球藻、团藻等一系列植物中,可以明显看到有性生殖进化过程,除团藻目以外,同样,还可以在丝藻目、管藻目中出现,这就表现出各个类群是各自独立地完成着有性生殖的进化。
比较高级的低等植物都是营异配卵式生殖,雄性生殖器官称为精子囊,雌性生殖器官称为卵囊,只有少数褐藻植物开始具有多室的配子囊结构。
高等植物的有性生殖器官都是多细胞结构,苔藓植物和蕨类植物的雌性生殖器官称为颈卵器,雄性生殖器官称精子器。在苔藓植物中,颈卵器和精子器最为发达,但随着类群越来越进化,有性生殖器官则变得越来越简化,到裸子植物仅有部分种类还保留着颈卵器,被子植物以胚囊和花粉管来代替精子器和颈卵器,从而完全摆脱了受精时需要水的条件。
关于有性生殖的起源问题,至今尚未完全解决。但可以发现植物体的细胞不经过分裂和转化,直接变为配子相结合的现象,如接合藻植物普遍如此;有些单细胞鞭毛植物,是以与一般的细胞完全相同的细胞结合的方式,进行有性生殖的,如衣藻属和丝藻属的游动孢子和配子,在形状、大小、结构等各方面都可以完全相同,在形态上没有区别,正常的配子在适当条件下,也可以单独发育成新的植物体。另一方面,正常的孢子也可能作为配子进行配合。这些事实说明,配子和孢子没有绝对界线,也就是说,有性生殖可能起源于无性生殖。
四、植物对陆地生活的适应
古生代以前,地球上一片汪洋,最原始的植物就是在这里产生和生活的,在20多亿年的漫长岁月里,它们在形态、结构、代谢和繁殖等方面,巴和水生环境相适应,并演化成形形色色的水生植物类群。到志留纪末期,整个地球发生了沧海桑田的大变动,陆地逐渐上升成为沼泽,海域逐渐缩小,不少生存在滨海或浅海潮汐地带的藻类,在适应新的环境过程中朝着各方面发展。某些藻类的后裔,逐渐加强其孢子体有利于陆生生活环境的变异性能,终于舍水登陆,产生了最早的以裸蕨植物为代表的第一批陆生植物。但是,陆地的生活环境显然和水里完全不同。首先登陆的裸蕨,它们在陆地生活中,摄取阳光和空气比在水中容易,这对于植物的生长发育非常有利。但是,在陆地生活易受到干旱的威胁,养料的摄取也不像整个身躯浸沉在水里那样容易,同时对如何支撑植物体直立在地上,也发生了很多困难,尤其是原来植物体的周围,赖以繁殖的水域条件已不存在。能否解决这些问题,关系到新生的陆生植物生死存亡的大问题,因而新生陆生植物的躯体,必须有一个适应和变异的过程。裸蕨植物能从水域到陆地生长,这是一个巨大的飞跃,在植物界的发展史上,标志着一个重要的里程碑。
裸蕨植物是形态结构简单,种类庞杂的一群植物,其中最有代表性的种类为莱尼蕨属。它虽然没有像一般高等植物那样有真正的根、茎和叶的分化,但是,它的根状茎和气生茎已出现了原始的维管组织,这就不仅有利于水和养料的吸收和运输,同时,也加强了植物体的支撑和固着作用;它的气生茎表面生有角质层和气孔,可以调剂水分的蒸腾;孢子囊大都生在枝的顶端,并且产生具有坚韧外壁的孢子,以利于孢子的传播和保护。凡此种种,裸蕨类植物已初步具有了适应多变的陆生环境的条件。但是,到泥盆纪末期,发生地壳的大变动,陆地进一步上升,气候变得更加干旱,裸蕨植物已不能再适应改变了的新环境,而趋向绝灭,盛极一时的裸蕨世界,让位于分化更完善、更能适应陆地生长的其他维管植物了。
维管植物的进化和发展,是向孢子体占优势的方向前进的。由于无性世代能够较好地适应陆生生活,孢子体得到充分的发展和分化,在形态、结构和功能上,都保证了陆生生活所必须的条件。它们的配子体在适应陆地生活上,受到了一定的限制,苔藓植物是朝着配子体发达的方向发展,这也是苔藓植物不能在植被中占重要地位的原因。
蕨类植物的孢子体,已具备各种适应陆地生活的组织机构,虽然这些组织还是初级的类型,但它已经能够在陆地上生长和发育,不过它的配子体还不能完全适应陆地生活,特别在受精作用时,还不能脱离水的环境。在蕨类植物中,有的出现了大、小孢子的分化,这两种孢子不仅形状、大小、结构不同,而且发育的前途也完全不同。大孢子发育成雌配子体,小孢子发育成雄配子体,而且雌、雄配子体终生不脱离孢子壁的保护,最后导致种子植物的出现。种子植物的配子体更为退化,几乎终身寄生在孢子体上,受精时,借花粉管将精子送入胚囊,与卵受精,从而克服了有性生殖时不能脱离水的缺点。被子植物又有了更进一步的发展,所以,它在如此复杂多变的陆地环境中,占绝对优势,这是发展的必然趋势。
五、生活史的类型及其演化
原核生物的生殖方式是细胞分裂和营养繁殖,所以它们的生活史非常简单。在真核生物发生之后的一定阶段,才出现了有性生殖。凡是进行有性生殖的植物,在它们的生活史中都有配子的配合过程和进行减数分裂过程。也就是说在它们生活史中存在着双相(2n)和单相(n)的核相的交替。进行有性生殖的植物,根据其减数分裂进行的时期,可分为3种类型:
1.减数分裂是在合子萌发前进行(合子减数分裂)。这种类型在藻类植物中相当普遍,如绿藻中的衣藻、团藻、丝藻、轮藻都属于这一类型。以衣藻为例:植物体是单倍的,除了有性生殖方式外,还有无性生殖或营养繁殖。有性生殖时,两个配子互相配合成合子,合子一萌发就进行减数分裂,形成单倍的孢子。在植物体的整个生活史中,合子实际上就成了唯一的二倍体阶段,而不再出现第二个二倍体的植物体。所以,这一类植物只存在着单倍的和二倍的核相的交替,而没有世代交替。
2.减数分裂在配子产生时进行(配子减数分裂)。这种类型在绿藻门中的管藻目和褐藻门中的无孢子纲植物,以及多种硅藻中普遍存在。以褐藻门的鹿角菜为例:这些植物的营养体为二倍的,减数分裂在配子产生前进行,合子萌发又成为二倍的植物体。在它们的生活史中,配子是生活史中唯一的单倍体阶段,没有再出现第二个单倍体植物体,所以也没有世代交替出现。动物和人类也是属于这个类型的。
3.减数分裂在二倍体的植物体产生孢子时进行(居间减数分裂)。绿藻中的石莼、浒苔,褐藻门除了无孢子纲植物外的各种类,红藻门的石花菜、多管藻,以及所有的高等植物全都属于这一类型。在这一类型的植物中,是二倍体的孢子体在产生孢子时进行减数分裂,孢子萌发成为单倍体的配子体,配子体所产生的精子和卵,结合成二倍的合子,分裂后形成胚,胚发育仍为二倍体的孢子体。所以这一类型的生活史中有产生孢子的二倍体植物,也有能产生配子的单倍体植物,而且在整个生活史中,二者是相互交替出现的。
在居间减数分裂类型中,有同型世代和异型世代之分,同型世代的像石莼、水云;异型世代又有配子体世代占优势的和孢子体世代占优势的两种,前者像苔藓植物,后者如褐藻中的海带以及所有维管植物的生活史。在低等植物中,除配子体有性别之外,孢子本身在形态上完全相同,但在蕨类植物的卷柏属和水生真蕨植物,孢子的形态、大小和生理机能都不相同,小孢子发育成雄配子体,大孢子发育成为雌配子体,配子体已更进一步简化,而且雌、雄配子体都是在孢子壁内萌发和发育的,并始终不脱离孢子壁的保护,在植物生活史上,这又是新的发展。
总之,植物生活史类型的演化过程,是随着整个植物界的进化而发展着,它经历了由简单到复杂,由低级到高级的演化过程。像细菌和蓝藻等原核植物是没有世代交替,也没有核相的交替的。到真核生物出现以后,才开始出现了有性生殖的核相交替,随后再出现世代交替。世代交替中,以居间减数分裂类型在植物界中最为高等,其中尤以不等世代交替中的孢子体世代越占优势,则越是进化。
六、高等植物营养体和孢子叶的发展与分化
在裸蕨植物没有发现以前,高等植物的根、茎、叶等器官如何发生和起源问题,很早就有了争论,但大都建立在缺乏充足的科学论据基础上,仍是模糊不清。譬如,有人认为最早出现的是叶子,茎和根是后来在进化过程中产生的。也有人认为被子植物的根、茎、叶、花、果实和种子等器官,在植物界和植物体的出现是同时产生的。自从发现了裸蕨植物以后,大大扩大了我们对这方面的知识,认识到最早的原始维管植物,大都无根,无叶,只有1个具二叉分枝的和能独立生活的体轴。这表明茎轴是原始维管植物最先出现的器官,并且能代行光合作用;其后,茎轴上发生了叶,才有茎、叶分化;最后出现的是根。
20世纪初,裸蕨植物的化石陆续发现,顶枝学说(Telome Theory)逐步得到充实,并且得到较为普遍的承认。顶枝学说认为,原始维管植物中,无叶的植物体(茎轴)是由顶枝(telome)构成的;顶枝是二叉分枝的轴的顶端部分,具有孢子囊或不具孢子囊,它的形体与莱尼蕨属(图5-52)相似,若干顶枝共同联合组成顶枝束,顶枝束的基部也有二叉分枝的部分,其表面生有假根。
关于叶的起源问题,顶枝学说认为,无论大型叶或是小型叶,都是由顶枝演变而来的,大型叶是由多数顶枝联合并且变扁而形成的(图6-1)。小型叶则是由单个顶枝扁化而成(图6-2,A-E)。

关于小型叶的来源,另一种是突出学说(Enation Theory),有完全不同的解释。突出学说认为,石松类的小型叶起源于茎轴表面的突出体,叶脉是后来才发生的(图6-2,F-J)。古植物学的有些资料,如裸蕨属植物(图5-55)的刺状突出物和星木属(图5-56)叶的结构情况,是和这种学说的观点基本相符合的。

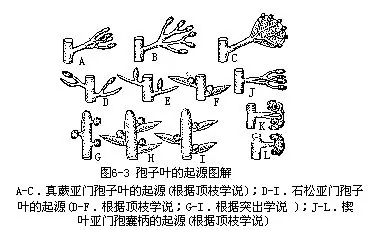
关于孢子叶的起源问题,羽叶植物类叶子的来源,根据顶枝学说,通常认为这是由1个能育顶枝束中的分子侧面结合的结果(图6-3,A—C),原来生在枝端的孢子囊成为生在孢子叶的边缘。
石松亚门植物孢子叶的起源,有两种说法。顶枝学说认为,孢子叶不育部分和孢子囊本身是同一个顶枝束发展而来的(图6-3,D—F);突出学说则认为,首先有孢子囊,后来形成突出体,突出体再和孢子囊组成孢子叶(图6-3,G-I)。
楔叶亚门植物的盾形孢囊柄,一般认为是特化了的孢子叶。根据顶枝学说,是由具孢子囊的顶枝束,经过顶部的弯曲和并联而形成的(图6-3,J-L)。
根的起源问题,由于化石资料不足,研究得还不够,有人认为是从裸蕨目的根状茎转变而成,是星木属的拟茎部分向下伸出的那种根状分枝发展而来的;也有人认为,根是从裸蕨植物地下部分的假根转变而来;还有人认为,根是后来产生的新结构。
七、植物的个体发育与系统发育
植物分类的基本单位是种,每个种又是由无数的个体组成。每一个体都有发生、生长、发育,以至成熟的过程,这一过程便称为个体发育(ontogeny)。在植物发育过程中,除外部形态发生一系列的变化外,其内部结构也随之出现组织分化,直到这一分化过程完全成熟,才达到比较完善的地步。
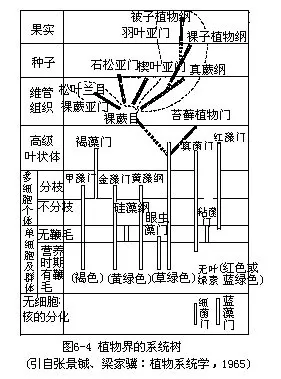
所谓系统发育(phylogeny)是与个体发育相对而言的,它是指某一个类群的形成和发展的过程。大类群有大类群的发展史,小类群有小类群的发展史,从大的方面看,如果考察整个植物界的发生与发展,便称之为植物界的系统发育。同样,也可以考察某个门、纲、目、科、属的系统发育,甚至在一个包含较多种以下单位的种(亚种,变种)中,也存在种的系统发育问题。例如,在绿藻门中有各种类型的植物,有单细胞的(其中又包括有鞭毛能活动和无鞭毛不能活动的两种类型),有群体的,有丝状体型的(其中又可分为分枝的和不分枝的),片状体型的……等。各种类型之间在进化上有何联系?哪种类型较为原始?哪种类型较为进化?何者低级?何者高级?对这类问题的探讨就是探讨绿藻门的系统发育。种是分类的基本单位,但在种之下又有亚种、变种、变型,这说明在一个种的范围内,也有变化和发展,这就是种的系统发育。同样道理,纲、目、科、属,各个分类等级均有其系统发育(图6-4)。
个体发育与系统发育,是推动生物进化的两种不可分割的过程,系统发育建立在个体发育的基础上,而个体发育又是系统发育的环节。在个体发育过程中,新一代的个体,既有继承上一代个体特性的遗传性,又有不同于上一代的变异性,种瓜得瓜,种豆得豆,这是遗传性决定的。但世上找不到两个完全相同的个体,即使是挛生兄弟,也有微小的差别,这就是变异性。自然界对新一代无数的大同而小异的个体进行选择,使有利于种族生存的变异得以巩固和发展,由量的积累而到质的飞跃,这就产生出了新的物种。只要生命物质存在一天,这一过程就永远不会休止。
在植物界中,任何高等植物的个体发育,都是从1个受精卵细胞开始的,这相当于进化过程中的单细胞阶段,由此细胞经过一系列的横分裂成为短小的丝状体,相当于丝状藻阶段,继之山现了多方面的分裂,外形趋于复杂化,这与片状藻和分枝丝状藻阶段大体相符,最后内部出现组织分化,出现了维管组织,这又象征着进入了维管植物的阶段,重演现象的发现,在进化论与有神论进行激烈争论的19世纪,为进化论提供了有力的佐证。
复习思考题
1.植物界从其出现到如今经过了哪几个主要发展阶段?这与地球环境的变迁有何关系。
2.试述植物界由单细胞到群体、到多细胞体的发展过程。
3.植物有性生殖经历了怎样的发展过程?
4.试从生态适应方面论述植物界的发展过程。
5.植物的生活史有哪几种主要类型?它们是怎样演化的。
6.何谓顶枝学说?它是怎样解释植物营养体和孢子叶进化的
7.何谓个体发育?何谓系统发育?两者之间存在什么关系