第一节 裸子植物的一般特征
裸子植物在植物界中的地位,介于蕨类植物和被子植物之间。它是保留着颈卵器,具有维管束,能产生种子的一类高等植物。
-
一、裸子植物的主要特征
(一)孢子体发达
裸子植物的孢子体特别发达,都是多年生木本植物,大多数为单轴分枝的高大乔木,枝条常有长枝和短枝之分。网状中柱,并生型维管束,具有形成层和次生生长;木质部大多数只有管胞,极少数有导管;韧皮部中无伴胞。叶多为针形、条形或鳞形,极少数为扁平的阔叶;叶在长枝上螺旋状排列,在短枝上簇生枝顶;叶常有明显的、多条排列成浅色的气孔带(stomatalband)。根有强大的主根。
(二)胚珠裸露
孢子叶(sporophyll)大多数聚生成球果状(strobiliform),称孢子叶球(strobilus)。孢子叶球单生或多个聚生成各种球序,通常都是单性,同株或异株;小孢子叶(雄蕊)聚生成小孢子叶球(雄球花staminate strobilus),每个小孢子叶下面生有贮满小孢子(花粉)的小孢子囊(花粉囊);大孢子叶(心皮)丛生或聚生成大孢子叶球(雌球花femal cone),胚珠裸露,不为大孢子叶所形成的心皮所包被,大孢子叶常变态为珠鳞(松柏类)、珠领(银杏)、珠托(红豆杉)、套被(罗汉松)和羽状大孢子叶(铁树)。而被子植物的胚珠则被心皮所包被,这是被子植物与裸子植物的重要区别。
(三)具有颈卵器的构造
裸子植物除百岁兰属(Welwitschia)、买麻藤属(Gnetum)外,具颈卵器。配子体完全寄生在孢子体上,雌配子体的近珠孔端产生颈卵器,但结构简单,埋藏于胚囊中,仅有2-4个颈壁细胞露在外面。颈卵器内有1个卵细胞和1个腹沟细胞,无颈沟细胞,比起蕨类植物的颈卵器更为退化。
(四)传粉时花粉直达胚珠
在被子植物中,花粉粒需先到柱头后萌发,形成花粉管,然后到达胚珠。而裸子植物则不同,花粉粒由风力(少数例外)传播,并经珠孔直接进入胚珠,在珠心上方萌发,形成花粉管,进达胚囊,使其内的精子与卵细胞受精。从传粉到受精这个过程,在裸子植物需经相当长的时间。有些种类在珠心的顶部具有花粉室(pollen chamber),以准备花粉粒在萌发前的逗留。
(五)具多胚现象
大多数裸子植物都具有多胚现象(polyembryony),这是由于1个雌配子体上的几个或多个颈卵器的卵细胞同时受精,形成多胚,称为简单多胚现象;或者由于1个受精卵,在发育过程中,胚原组织分裂为几个胚,这是裂生多胚现象(cleavage polyembryony)。
此外,花粉粒为单沟型,具气囊或缺如,无3沟、3孔沟或多孔的花粉粒等也是裸子植物的特征。
在裸子植物这一章中,有两套名词时常并用或混用:一套是在种子植物中习用的,如“花”、“雄蕊”、“心皮”等;一套是在蕨类植物中习用的,如“孢子叶球”、“小孢子叶”、“大孢子叶”等。这种情况的产生是有其历史原因,19世纪中叶以前,人们不知道种子植物的这些结构和蕨类植物的结构有系统发育上的联系,所以出现了这两套名词。1851年,德国植物学家荷夫马斯特(Hofmeister)将蕨类植物和种子植物的生活史完全贯通起来,人们才知道裸子植物的球花相当于蕨类植物的孢子叶球,前者是后者发展而来。
二、裸子植物的分类
裸子植物在植物分类系统中,通常作为1个自然类群,称为裸子植物门(Gymnosper-mae)。裸子植物门通常分为铁树纲(Cycadopsida)、银杏纲(Ginkgopsida)、松柏纲(球果纲)(Coniferopsida)、红豆杉纲(紫杉纲)(Taxopsida)及买麻藤纲(倪藤纲)(Gnetopsida)(盖子植物纲)(Chalmydospermopsida)5纲。
裸子植物发生发展的历史悠久,最初的裸子植物出现,约在34500万年前至39500万年之间的古生代泥盆纪,历经古生代的石炭纪、二叠纪,中生代的三叠纪、侏罗纪、白垩纪,新生代的第三纪、第四纪。从裸子植物发生到现在,地史气候经过多次重大变化,裸子植物种系也随之多次演变更替,老的种类相继灭绝,新的种类陆续演化出来,种类演替繁衍至今。现代的裸子植物有不少种类,是从约250万年前至6500万年之间的新生代第三纪出现的,又经过第四纪冰川时期保留下来。现代裸子植物的种类分属于5纲,9目,12科,71属,近800种。我国是裸子植物种类最多、资源最丰富的国家,有5纲,8目,11科,41属,236种。其中引种栽培1科,7属,51种。有不少是第三纪的孑遗植物,或称“活化石”植物。我国的裸子植物多为林业经营上的重要用材树种,也是纤维、树脂、单宁等原料树种,少数种类的枝叶、花粉、种子、根皮等可供药用。
第二节 铁树纲(苏铁纲)(Cycadopsida)
常绿木本植物,茎干粗壮,常不分枝。叶螺旋状排列,有鳞叶及营养叶,二者相互成环着生;鳞叶小,密被褐色毡毛;营养叶大,羽状深裂,集生于树干顶部。孢子叶球亦生于茎顶,雌雄异株。游动精子有多数纤毛。染色体:X=8、9、11、13。
铁树纲植物在古生代的末期(二叠纪)兴起,中生代的侏罗纪相当繁盛,以后逐渐趋于衰退,现存的仅有1目,1科,共9属,约110种,分布于南、北半球的热带及亚热带地区,其中4属产美洲、2属产非洲、2属产大洋洲、1属产东亚。我国仅有铁树属(Cycas),8种。
铁树科(Cycadaceae)的铁树(Cycas revoluta Thunb.)(图7-1,A)具柱状主干,常不分枝,顶端簇生大型羽状深裂的叶。茎中有发达的髓部和甚厚的皮层,网状中柱,内始式木质部,次生木质部的管胞具多列圆形的缘孔,形成层的活动期较短,后为由皮层相继发生的形成层环所代替。叶为一回羽状深裂,革质,坚硬,幼时拳卷,脱落后茎上残留有叶基。侧根具有特化的菌根,菌根内还有鱼腥藻属(Anabaena)共生。
铁树的大、小孢子叶异株。小孢子叶扁平,肉质,鳞片状窄楔形,具短柄,紧密地螺旋状排列成椭圆形的小孢子叶球,生于茎顶。每个小孢子叶下面生有许多由3-5个小孢子囊组成的小孢子囊群(图7-1, B、C)。小孢子囊是厚囊性发育,囊壁由多层细胞构成,借表皮细胞壁不均匀增厚而纵裂,散发小孢子。这是裸子植物中孢子囊机械组织构造与蕨类植物相似的唯一代表。小孢子多数,两侧对称,宽椭圆形,具1纵长的深沟。大孢子叶丛生于茎顶,密被淡黄色绒毛,上部羽状分裂,下部成狭长的柄,柄的两侧生有2—6枚胚珠。胚珠较大,直生,有1层珠被,珠心顶端有喙及花粉室,与珠孔相通,珠心内的胚囊发育有2—5个颈卵器(图7-2,A、B、C)。颈卵器位于珠孔下方,颈部短小,通常仅由2个细胞组成,受精的前几天,中央细胞的核分为2,下面一个变为卵核,上面1个是不发育的腹沟细胞,并很快消失。
小孢子萌发,形成具有3个核的雄配子体(图7-2,D),即基部1个原叶体细胞(营养细胞),此细胞不再分裂;上面的1个细胞再分裂一次成为1个管细胞(吸器细胞)及l个生殖细胞,并以3个细胞状态从小孢子囊中散出,随风传播到珠孔上。由珠孔溢出1滴液体,名为传粉滴(pollination drop),雄配子体随着液滴的干涸而被吸入花粉室。随后生殖细胞分裂为2,大的叫体细胞,小的叫柄细胞。体细胞又分裂为2个精细胞,成熟的精子为陀螺形,端有纤毛,能游动,长可达0.3毫米,是生物界中最大的精子。管细胞的主要功用不是输送精子,而是吸取养料,当先端生长,伸至颈卵器旁时即炸裂,2个游动精子进入颈卵器,1个与卵结合,形成合子,另1个消失。
合子在长时间内重复进行游离核分裂,形成未分化的原胚。原胚分化缓慢,基部一些细胞伸长形成胚柄,原胚的末端则分化发育成胚。种子成熟时,胚已发育成具有2片子叶和稍指向珠孔的胚根的大形圆柱体,并陷于充满营养物质的雌配子体中,雌配子体此时又称为胚乳。珠被变成种皮,分为3层:外层肉质甚厚,中层为石细胞所成的硬壳,内层为薄纸质。种子无休眠期,萌发时,根由珠孔穿出,子叶则永留种子中吸取养料。


铁树栽培极为广泛,为优美的观赏树种;茎内髓部富含淀粉,可供食用;种子含油和丰富的淀粉,微有毒,供食用和药用,有治痢疾、止咳及止血之效。
第三节 银杏纲(Ginkgopsida)
落叶乔木,枝条有长、短枝之分。叶扇形,先端2裂或波状缺刻,具分叉的脉序,在长枝上螺旋状散生,在短枝上簇生。球花单性,雌雄异株,精子具多纤毛。种子核果状,具3层种皮,胚乳丰富。
本纲现仅残存1目,1科,1属,1种,为我国特产,国内外栽培很广。染色体: X=12。
银杏科(Ginkgoaceae)的银杏(Ginkgo biloba L.)(图7-3,A)为落叶乔木,树干高大,枝分顶生营养性长枝和侧生生殖性短枝。两种枝的解剖构造亦极不同:长枝髓小,皮层薄,木质部甚厚;短枝则正相反,髓大,皮层厚,木质部甚窄。网状中柱,内始式木质部,原生木质部仅有螺纹管胞,后生木质部为孔纹管胞,次生木质部则由具缘纹孔的管胞组成,年轮明显。各种器官内均有分泌腔。叶扇形,有柄,长枝上的叶大都具2裂,短枝上的叶常具波状缺刻。
银杏雌雄异株,营养体雌、雄株的主要区别有:雄株树冠狭圆锥形,雌株阔圆锥形;雄株长枝斜上伸展,大枝基部具乳状突瘤,雌株长枝开展和下垂,无乳状突;雄株叶柄横切无树脂隙,雌株具树脂隙;雄株实生苗幼根直伸,无乳状突,雌株稍屈曲,具乳状突;雄株苗木形大、干细、横枝少,叶大而多裂,雌株形小、干粗、横枝多、叶小裂少。
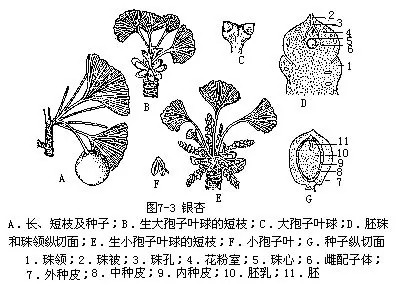
球花单性。小孢子叶球呈柔荑花序状,生于短枝顶端的鳞片腋内。小孢子叶有1短柄,柄端有由2个(稀为3—4个,或甚至7个)小孢子囊组成的悬垂的小孢子囊群。大孢子叶球很简单,通常仅有1长柄,柄端有2个环形的大孢子叶,称为珠领(collar),也叫珠座,大孢子叶上各生1个直生胚珠,但通常只有1个成熟,偶有若干个胚珠,这是一种返祖现象。珠被1层,珠心中央凹陷为花粉室。雌配子体发育极似铁树,不同的是珠被发育时含有叶绿素,并有明显的腹沟细胞。雄配子体的发育和受精过程等,也都与铁树相似。具纤毛的游动精子是受精时需水的遗迹,这是铁树与银杏所具有的原始性状。
胚的发育步骤和成熟种子,也与铁树相似,不过胚柄不强烈伸长,胚的体积较小。种子近球形,熟时黄色,外被白粉,种皮分化为3层(图7-3,G):外种皮厚,肉质,并含有油脂及芳香物质;中种皮白色,骨质,具2—3纵脊;内种皮红色,纸质。胚乳肉质。子叶2,不出土。
银杏为著名的孑遗植物,为我国特产,现广泛栽培于世界各地,仅浙江西天目山有野生状态的树木,生于海拔 500—1000米。栽培的银杏有数百年或千年以上的老树,树形优美,春季叶色嫩绿,秋季鲜黄,颇美观,可做行道树及园林绿化的珍贵树种。木材优良,可供建筑、雕刻、绘图板、家具等用材。种仁(白果)供食用(多食易中毒)及药用,入药有润肺、止咳、强壮等功效,叶供药用和制杀虫剂,树皮含单宁。
第四节 松柏纲(球果纲)(Coniferopsida)
一、松柏纲的主要特征
常绿或落叶乔木,稀为灌木,茎多分枝,常有长、短枝之分;茎的髓部小,次生木质部发达,由管胞组成,无导管,具树脂道(resin duct)。叶单生或成束,针形、鳞形、钻形、条形或刺形,螺旋着生或交互对生或轮生,叶的表皮通常具较厚的角质层及下陷的气孔。孢子叶球单性,同株或异株,孢子叶常排列成球果状。小孢子有气囊或无气囊,精子无鞭毛。球果的种鳞与苞鳞离生(仅基部合生)、半合生(顶端分离)及完全合生。种子有翅或无翅,胚乳丰富,子叶2—10枚。松柏纲植物因叶子多为针形,故称为针叶树或针叶植物;又因孢子叶常排成球果状,也称为球果植物。
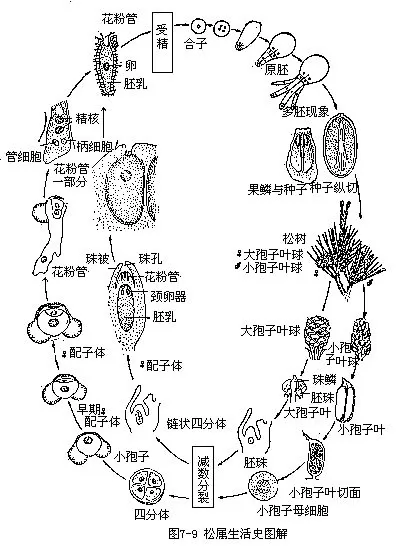
二、松柏纲植物的生活史
以最普遍而研究最详尽的松科(Pinaceae)的松属(Pinus)为代表,介绍如下。
(一)孢子体
松属的孢子体为高大多年生常绿乔木,单轴分枝,主干直立,旁枝轮生,具长枝和短枝。网状中柱,90—95%由管胞组成,树脂道约占1%,木射线约占6%。长枝上生鳞叶,腋内生短枝,短枝极短,顶生1束针形叶,每束通常2、3、5个叶,基部常有薄膜状的叶鞘8—12枚(由芽鳞变成)包围,叶内有1或2条维管束和几个树脂道。
孢子叶球单性,同株。小孢子叶球排列如穗状,生在每年新生的长枝条基部,由鳞片叶腋中生出。每个小孢子叶球有1个纵轴,纵轴上螺旋状排列着小孢子叶,小孢子叶的背面(远轴面)有1对长形的小孢子囊。小孢子囊内的小孢子母细胞,经过两次的连续分裂(其中一次为减数分裂),形成4个小孢子(花粉粒)。小孢子有2层壁,外壁向两侧突出成气囊,能使小孢子在空气中飘浮,便于风力传播。
大孢子叶球1个或数个着生于每年新枝的近顶部,初生时呈红色或紫色,以后变绿,成熟时为褐色。大孢子叶球是由大孢子叶构成的,大孢子叶也是螺旋状排列在纵轴上的,但它们不是简单的孢子叶,而是由两部分组成:下面较小的薄片称为苞鳞(bract);上面较大而顶部肥厚的部分称为珠鳞(ovuliferous scale),也叫果鳞或种鳞,一般认为珠鳞是大孢子叶,苞鳞是失去生殖能力的大孢子叶。在松科各属植物苞鳞和珠鳞是完全分离的,每1珠鳞的基部近轴画着生2个胚珠,胚珠由1层珠被和珠心组成,珠被包围着珠心,形成珠孔。珠心即大孢子囊,中间有1个细胞发育成大孢子母细胞,经过两次连续分裂(其中一次是减数分裂),形成4个大孢子,排列成1列称为“链状四分体”。通常只有合点端的1个大孢子发育成雌配子体,其余3个退化。

(二)雄配子体
雄配子体是1个大为减退了的结构,只由少数几个细胞构成(图7-4,A、B、C、D)。小孢子(单核时期的花粉粒)是雄配子体的第一个细胞,小孢子在小孢子囊内萌发,细胞分裂为2,其中较小的1个是第一个原叶体细胞(营养细胞),另1个大的叫胚性细胞,胚性细胞再分裂为2,即第二原叶细胞及精子器原始细胞(中央细胞),精子器原始细胞再分裂为2,形成管细胞和生殖细胞。成熟的雄配子体有4个细胞:2个退化原叶体细胞、1个管细胞和1个生殖细胞。
(三)雌配子体
由大孢子发育而成。因此,大孢子是雌配子体的第一细胞,它在大孢子囊(珠心)内萌发,进行游离核分裂,形成具16—32个游离核,不形成细胞壁。雌配子体的四周具1薄层细胞质,中央为1个大液泡,游离核多少均匀分布于细胞质中,当冬季到来时,雌配子体即进入休眠期。翌年春天,雌配子体重新开始活跃起来,游离核继续分裂,主要表现游离核的数目显著增加,体积增大。以后雌配子体内的游离核周围开始形成细胞壁,这时珠孔端有些细胞明显膨大,成为颈卵器的原始细胞。之后,原始细胞进行一系列的分裂,形成几个颈卵器,成熟的雌配子体包含2—7个颈卵器和大量的胚乳(图7-5)。

(四)传粉与受精
传粉在晚春进行,此时大孢子叶球轴稍为伸长,使幼嫩的苞鳞及珠鳞略为张开。同时,小孢子囊背面裂开一条直缝,处于雄配子体阶段的花粉粒,借风力传播,飘落在由珠孔溢出的传粉滴中,并随液体的干涸而被吸入珠孔。这时大孢子叶球的珠鳞又继续闭合。雄配子体中的生殖细胞分裂为2,形成1个柄细胞及1个体细胞,而管细胞则开始伸长,迅速长出花粉管(图7-4,E)。但这时大孢子尚未形成雌配子体,花粉管进入珠心相当距离后,即暂时停止伸长,直到第二年春季或夏季颈卵器分化形成后,花粉管才再继续伸长,此时体细胞再分裂形成2个精子(不动精子)。受精作用通常是在传粉以后13个月才进行,即传粉在第一年的春季,受精在第二年夏季。这时大孢子叶球已长大并达到或将达到其最大体积,颈卵器已完全发育。当花粉管伸长至颈卵器,破坏颈细胞到达卵细胞处,其先端随即破裂,2个精子、管细胞及柄细胞都一起流入卵细胞的细胞质中,其中1个具功能精子随即向中央移动,并接近卵核,最后与卵核结合形成受精卵,这个过程称受精。
(五)种子
松属的胚胎发育过程,颇复杂,具明显的阶段性,通常可分原胚阶段、胚胎选择阶段、胚的器官和组织分化阶段、胚的成熟阶段。但这些阶段是按顺序连续发育的,是相互联系和相互制约的。
1.原胚阶段 从受精卵分裂开始到细胞型原胚的形成,先后经过游离核的分裂、细胞壁的产生和原胚的形成(图7-6)。受精卵接连进行3次游离核的分裂,形成8个游离核,这8个游离核在颈卵器基部排成上、下两层,每层4个,细胞壁即在此时形成,但上层4个细胞的上部不形成胞壁,使这些细胞的细胞质与卵细胞质相通,称为开放层;下层4个细胞称为初生胚细胞层。接着开放层和初生胚细胞层各自再分裂1次,形成4层,分别称为上层、莲座层、胚柄层(初生胚柄层)和胚细胞层,组成原胚(proembryo)。

2.胚胎选择阶段 胚柄系统的发育和多胚现象的产生是这个阶段的主要特征。原胚的4层细胞从上到下,第一层(上层),初期有吸收作用,不久即解体;第二层莲座层,分裂数次之后消失;第三层胚柄层,它的4个细胞称为初生胚柄(primary suspensor),不再分裂,但伸长;第四层胚细胞层的胚细胞,在胚柄细胞继续延长的同时,紧接着后面的胚细胞进行分裂并伸长,称为次生胚柄(secondary suspensor),由于胚柄和次生胚柄(胚管)迅速伸长,形成多回卷曲的胚柄系统。而胚细胞层的最前端的细胞发育成胚的本身,但它们不组成1个胚,而在纵面彼此分离,各个单独发育成胚,称为多胚现象。常见的多胚现象有两种:一种是简单多胚现象(图7-7,C),即在同一个胚珠内有2个以上的颈卵器的卵细胞,可以同时受精,因而在胚胎发育的早期,可以产生2个以上的原胚;另一种是裂生多胚现象(图7-7,D),即由1个受精卵形成的4个胚细胞,分别单独发育成为4个幼胚。在胚胎发育过程中,通过胚胎选择,通常只有1个(很少2个或更多)幼胚正常分化、发育,成为种子中成熟的胚。
大多数松柏类植物具有2个以上的颈卵器,所以简单多胚现象是普遍发生的,至于裂生多胚现象,则仅限于松柏类植物的几个属可见。
3.胚的器官和组织分化阶段 胚在进一步的发育中成为1个伸长的圆柱体。这个圆柱体的近轴区(基部)同胚柄系统相接,主要是横分裂,细胞略大,形成较规则的行列,进而发育成根端和根冠组织;而在远轴区内,细胞分裂似无特定的方向,细胞较小,由这些细胞进一步分化,最后分裂出下胚轴、胚芽和子叶。
4.胚的成熟阶段 成熟的胚包括胚根、胚轴(胚茎)、胚芽和子叶(通常7-10枚)。包围此胚的雌配子体(胚乳)继续生长,最后珠心仅遗留一薄层。珠被发育成种皮,种皮分为3层:外层肉质(不发达)、中层石质、内层纸质。
胚、胚乳、种皮构成种子(图7-8)。裸子植物的种子是由3个世代的产物组成的,即胚是新的孢子体世代(2n);胚乳是雌配子体世代(n);种皮是老的孢子体(2n)。受精后,大孢子叶球继续发育,珠鳞木质化而成为种鳞,种鳞顶端扩大露出的部分为鳞盾,鳞盾中部有隆起或凹陷的部分为鳞脐,珠鳞的部分表皮分离出来形成种子的附属物即翅,以利风力传播。种子萌发时,主根先经珠孔伸出种皮,并很快产生侧根,初时子叶留在种子内,从胚乳中吸取养料,随着胚轴和子叶的不断发展,种皮破裂,子叶露

出,而随着茎顶端的生长,产生新的植物体。松属生活史图解见(图7-9)。
三、分类及代表植物
松柏纲植物是现代裸子植物中数目最多、分布最广的类群。现代松柏纲植物有44属,约400余种,隶属于4科,即松科、杉科、柏科和南洋杉科(Araucariaceae),它们分布于南、北两半球,以北半球温带、寒温带的高山地带最为普遍。我国是松柏纲植物最古老的起源地,也是松柏植物最丰富的国家,并富有特有的属、种和第三纪孑遗植物,有3科,23属,约150种,为国产裸子植物中种类最多,经济价值最大的1个纲,分布几遍全国。另引入栽培1科,7属,50种,多为庭园绿化及造林树种。
(一)松科(Pinaceae)
乔木,稀灌木,大多数常绿。叶条形或针形:条形叶扁平,稀呈四棱形,在长枝上螺旋状散生,在短枝上簇生;针形叶常2-5针成束,着生于极度退化的短枝顶端,基部包有叶鞘。孢子叶球单性同株,小孢子叶球,具多数螺旋状着生的小孢子叶,每个小孢子叶有2个小孢子囊,小孢子多数有气囊;大孢子叶球,由多数螺旋状着生的珠鳞与苞鳞所组成,每珠鳞的腹面(上面)具两枚倒生的胚珠,背面(下面)的苞鳞与珠鳞分离(仅基部结合),花后珠鳞增大发育成种鳞。球果直立或下垂。种子通常有翅;胚具2-16枚子叶。染色体:X=12、13、22。
本科是松柏纲植物中最大而且在经济上又是重要的1科,有10属,约230余种,多产于北半球。我国有10属,113种(包括引种栽培24种),分布遍于全国,绝大多数都是森林树种和用材树种,以及许多还是特有属和孑遗植物。
1.冷杉属(Abies) 枝具圆形而微凹的叶痕。叶条形,上面中脉凹下。叶内具2个(稀4-12)树脂道。球果直立,当年成熟,种鳞和种子一同脱落。约50种,分布于亚洲、欧洲、北美及非洲北部高山地带。我国有19种,分布于东北、华北、西北及浙江、台湾等省区的高山地带,常组成大面积的纯林,用途很广,为今后开发利用的主要森林资源,亦为森林更新的主要树种。常见的有臭冷杉[A.nephrolepis(Trautv.)Maxim.],一年生枝有毛,树脂道中生,产于东北、华北。冷杉[A.fabri(Mast.)Craib],一年生枝无毛,树脂道边生,为我国特有树种,产于四川。百山祖冷杉(A.beshanzuensis M.H.Wu),为我国新近发现的稀有珍贵树种,产于浙江南部百山祖山。
2.银杉属(Cathaya)(图7-10) 枝分长、短枝。叶条形扁平,上面中脉凹陷,在枝节间的顶端排列紧密成簇生状,在其下则排列疏散。球果腋生,初直立后下垂,种鳞远较苞鳞大,宿存。仅银杉(C.argyrophylla Chun et Kuang)1种,分布于广西及四川,为我国特产的稀有树种,材质优良,供建筑、家具等用材。
3.云杉属(Picea) 小枝有显著隆起的叶枕。叶四棱状条形或条形,无柄,四面有气孔线或仅上面有气孔线。孢子叶球单性同株。球果下垂。种鳞宿存,苞鳞短小。约40种,分布于北半球。我国有16种,另引种栽培2种,产于东北、华北、西北、西南等省区及台湾省的高山地带,常组成大面积的自然林,为我国主要的林业资源之一。材质优良,纹理细致,结构紧密,有弹性,可供建筑、飞机、舟车、家具等用材,树干可取松脂,树皮可制栲胶。常见的有云杉(P.asperata Mast.),一年生枝褐黄色或淡黄褐色,多少有白粉,叶横切面为四棱形,球果长通常6—10厘米,为我国特有树种,产陕西、甘肃及四川。鱼鳞云杉[P.jezoensis var.microsperma(Lindl.)Cheng et L.K.Fu],叶横切面扁平。球果长4—6厘米,产我国东北。
4.落叶松属(Larix) 落叶乔木,枝有长枝和距状短枝。叶条状扁平,在长枝上螺旋状散生,在短枝上呈簇生状。孢子叶球单生于短枝顶端,球果直立,当年成熟,种鳞革质,宿存。种子上部具膜质长翅。约18种,分布于北半球的亚洲、欧洲及北美洲的温带高山与寒温带、寒带地区。我国产10种,广布于东北、华北、西北、西南等省区,常组成纯林。材质坚韧,结构细致,抗腐性强,可供建筑、桥梁、舟车、家具等用材,树干可提取树脂,树皮可提取栲胶,亦可栽培作庭园树种。落叶松[L.gmelinii(Rupr.)Rupr.]小枝不下垂,球果卵圆形或椭圆形,苞鳞较种鳞为短,为我国东北林区的主要森林树种。红杉(L. Potaninii Batal.)小枝下垂,球果圆柱形或卵状圆柱形,苞鳞较种鳞为长,显著露出,为我国特有树种,产于甘肃、陕西及四川。
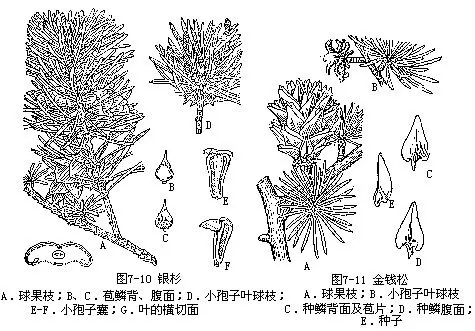
5.金钱松属(Pseudolarix) 落叶乔木,枝有长枝与短枝,短枝距状。叶条形,柔软、扁平,在长枝上螺旋状散生,距状短枝上呈簇生状,辐射平展呈圆盘形。小孢子叶球穗状,数个簇生。球果当年成熟,种鳞木质,成熟后脱落,种子有宽大种翅。仅有金钱松[P.amabilis(Nelson)Rehd.]1种(图7-11),为我国特有属种,产华东和华中及四川等省区。金钱松生长较快,喜生于温暖、多雨和土层深厚、肥沃、排水良好的酸性土山区。木材纹理通直,硬度适中,材质稍粗,性较脆,可作建筑、板材、家具、器具等用,树皮可提栲胶,入药(俗称土槿皮)可治顽癣和食积等症,种子可榨油。树姿优美,秋后叶呈金黄色,颇为美观。为优良有用材树种及庭园树种。
6.雪松属(Cedrus) 常绿乔木,枝有长枝和短枝。叶针形,坚硬,通常三棱形,或背脊明显而呈四棱形。球果第二年(稀三年)成熟,熟后种鳞从宿存的中轴上脱落,种子有宽大膜质的种翅。有4种,分布于非洲北部、亚洲西部及喜马拉雅山西部。我国有1种和引种栽培1种。雪松[C.deodara(Roxb.)Loud],材质坚实,致密而均匀,具香气,少翘裂而耐久用,可作建筑、桥梁、造船、家具等用。雪松终年常绿,树形美观,我国各大城市广泛栽培作庭园树种。
7.松属(Pinus) 常绿乔木,稀灌木。冬芽显著。叶有两型:鳞叶(原生叶)单生,螺旋状着生,在幼苗为扁平条形,后逐渐退化成膜质苞片状;针叶(次生叶)螺旋状着生,常2、3或5针一束,生于苞片状鳞叶的腋部,着生于不发育的短枝的顶端,每束针基部由8—12枝芽鳞组成的叶鞘所包,叶鞘脱落或宿存。孢子叶球单性同株,小孢子叶球多数,集生于新枝下部;大孢子叶球单生或2—4个生于新枝近顶端。球果第二年(稀第三年)秋季成熟,种鳞木质,宿存,种子上部具长翅。约80余种,广布于北半球。我国产22种,分布几遍全国,为我国森林中的主要树种。木材含有松脂,材质较硬或软,纹理直或斜,结构中至粗,可供建筑、枕木、桥梁、舟车、板材、家具及造纸等用材;树干可用以采脂;树皮、针叶、树根等可综合利用,制成各种化工产品;种子可榨油,有些可供食用;药用的松花粉、松节、松针及松节油是从各种松树采取和提取;多数种类为森林更新、造林、绿化及庭园树种。本属又可分2个亚属:即软松亚属或称单维管束亚属和硬松亚属或称双维管束亚属。
(1) 单维管束亚属(Strobus) 木材软,色淡。叶鞘早落,针叶基部的鳞叶不下延。叶内有1条维管束。常见的有:红松(P.koraiensis Sieb.et Zucc.),小枝密被黄褐色或红褐色柔毛,针叶5针一束,横切面近三角形,树脂道3个中生,种鳞的鳞脐顶生,产于我国东北。华山松(P. H mandii Franch.),小枝无毛,针叶5针一束,稀6—7针一束,横切面三角形,树脂道常3个中生或背面2个边生,腹面中生,种鳞鳞脐不明显,产于我国山西、陕西、河南、四川及云南等省山地。白皮松(P.bungeana Zucc.ex Endl.),幼树树皮光滑,灰绿色,老树皮成不规则的薄片块状脱落,呈淡褐灰色或灰色;小枝无毛;针叶3针一束,横切面扇状三角形,树脂道6—7个边生,稀背面角处1—2个中生;种鳞鳞脐背生,顶端有刺。为我国特有树种,产于山西、河南、陕西、甘肃、四川及内蒙等地。
(2) 双维管束亚属(Pinus) 木材硬,色深。叶鞘宿存,针叶基部的鳞叶下延。叶内有2条维管束。鳞脐背生,种子上部具长翅。常见的有:油松(P. tabulaeformis Carr.),小枝无毛,微被白粉,针叶2针一束,叶边缘有细锯齿,横切面半圆形,树脂道5—8个或更多,边生,鳞脐凸起有尖刺。为我国特有树种,产于华北、东北等地;马尾松(P.massoniana Lamb.),枝条无毛,针叶2针一束,稀3针一束,横切面半圆形,树脂道4—8个边生,鳞脐微凹无刺。主要分布于我国中部、长江流域以南各省区;黄山松(P.taiwanensis Hayata),枝条无毛,针叶2针一束,横切面半圆形,树脂道3—7个中生,鳞脐具短刺。为我国特有树种,产于台湾、安徽、福建、浙江及江西等省,海拔600—2800米山地;黑松(P.thunbergii Parl.),小枝淡褐黄色,无毛,冬芽银白色,针叶2针一束,粗硬,横切面半圆形,树脂道6—11个中生,鳞脐小而平,常有小棘。原产日本、朝鲜,我国辽宁及华东各省引种栽培,为造林和庭园观赏树种。
此外,本科植物我国产还有油杉属(Keteleeria)、黄杉属(Pseudotsuga)、铁杉属(Tsuga)等。
本科植物以具针形叶或条形叶,叶及种鳞螺旋状排列,种鳞与苞鳞离生,每种鳞具2粒种子等为特色。
(二)杉科(Taxodiaceae)
乔木。叶螺旋状排列,同一树上的叶同型或二型;孢子叶球单性同株,小孢子叶及珠鳞螺旋状排列(仅水杉的叶和小孢子叶、珠鳞对生),小孢子囊多于2个(常3—4个),小孢子无气囊,珠鳞与苞鳞多为半合生(仅顶端分离),珠鳞的腹面基部有2—9枚直立或倒生胚珠。球果当年成熟,种鳞(或苞鳞)扁平或盾形,木质或革质,能育种鳞有2—9粒种子,种子周围或两侧有窄翅。染色体:X=11、33。
本科有10属,16种,主要分布于北半球。我国产5属,7种,引入栽培4属,7种,分布于长江流域及秦岭以南各省区。
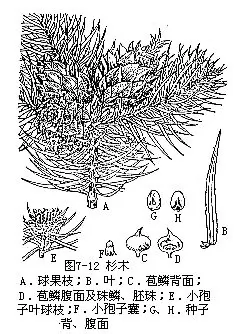
1.杉木属(Cunninghamia)常绿乔木。叶条状披针形,边缘有锯齿,螺旋状着生,叶的上、下两面均有气孔线。苞鳞与珠鳞的下部合生,螺旋状排列,苞鳞大,边缘有不规则细锯齿,珠鳞小,先端3裂,腹面基部生3枚胚珠。球果近球形或卵圆形,种子两侧具窄翅。共2种,为我国特产,分布于长江流域以南各省区及台湾省。杉木[C.lanceolata(Lamb.)Hook.] (图7-12),叶在主枝上辐射伸展,在侧枝上叶基扭转成二列状,叶缘有细锯齿,下面中脉两则各有10条白色气孔线,种子两侧边缘有窄翅。杉木为秦岭以南面积最大的人造林,生长快,经济价值高,材质优良,易于加工,可供建筑、桥梁、枕木、板材、家具等用材;树皮可提栲胶。台湾杉木(C.konishii Hayata),特产于我国台湾中部以北山区,为台湾省主要用材树种之一。
2.柳杉属(Cryptomeria)常绿乔木。叶钻形,螺旋状排列略成5行列,背腹隆起。小孢子叶球单生叶腋,大孢子叶球单生枝顶,每一珠鳞有2—5枚胚珠,苞片与珠鳞合生,仅先端分离,球果近球形,种子有极窄的翅。共2种分布于我国及日本。树干高大,材质轻软,纹理直,可供建筑、桥梁、板材及家具等用材,也是优美的园林树种。柳杉(C.fortunei Hooibrenk ex Otto et Dietr.),叶先端向内弯曲,种鳞较少,每1种鳞有2粒种子。为我国特有树种,产于浙江天目山、福建、江西等地。日本柳杉[C.japonica(Linn.f.)D.Don],叶先端微内曲,种鳞较多,每1种鳞有2—5粒种子。原产日本。
3.水松属(Glyptostrobus) 叶螺旋状着生,有3种类型:鳞形叶较厚,辐射伸展;条形叶薄,常排成2列状;条状钻形叶,辐射伸展成3列状。鳞形叶宿存,条形或条状钻形叶均于秋后连同侧生短枝一同脱落。球果直立,种鳞木质,上部边缘有6—10个三角状尖齿,能育种鳞有2粒种子,种子具向下生长的长翅。
在白垩纪和新生代广布于北半球,第四纪冰期后期,在欧洲、美洲、日本及其他各地均已绝灭,现存水松[G.pensilis(Lamb.) K.Koch]1种,仅产我国,分布于华南、西南,现各大城市均有栽培。材质轻软,纹理细,耐水湿,供建筑、家具等用材;根部材质轻松,浮力大,可作救生用品及瓶塞材料;树形优美,可作庭园树种。
4.落羽杉属(Taxodium) 小枝有2种,主枝宿存,侧生小枝冬季脱落。叶螺旋状排列,异型,钻形叶在主枝上斜上伸展,宿存;条形叶在侧生小枝上排成2列,冬季与枝一同脱落。小孢子叶球生于小枝顶端,常排成总状或圆锥状花序形。球果球形,角状,小尖头,能育种鳞有2粒种子,种子呈不规则三角形,有明显而锐利的棱脊。共有3种,原产北美及墨西哥,我国均有引种,作庭园和造林树种。落羽杉[T.distichum(Linn.)Rich.],叶条形,扁平,排成2列。原产北美。池杉(T.ascendens Brongn.),叶钻形,在枝上螺旋状伸展,原产北美,现已成为长江流域广大地区的优良造林树种。
5.水杉属(Metasequoia) 落叶乔木。小枝对生或近对生。条形叶交互对生,基部扭转排成2列,冬季与侧生小枝一同脱落。球果的种鳞盾形,木质,交互对生,能育种鳞有种子5—9粒,种子扁平,周围有窄翅。
本属在中生代白垩纪及新生代约有10种,曾广布于北美、中国、日本及苏联。第四纪冰期之后,几全部绝灭,现仅有水杉(M.glyptostroboides Hu et Cheng)(图7-13)1种,为我国特产,是稀有珍贵的孑遗植物,分布于四川石柱县、湖北利川县、湖南西北部等地,现各地普遍栽培。在国外,约有50多个国家和地区亦引种栽培。材质轻软、优良,可供建筑、板材及家具等用材;又因生长快,树姿挺直优美,为著名的庭园、绿化树种。
水杉在系统发育上与水松属、北美红杉属(红杉属,Sequoia)、巨杉属(世界爷属,Sequoia-dendron)和落羽杉属可能有亲缘关系,不过这4个属的叶及球果的种鳞都是互生的,而水杉的叶及种鳞是对生的,似又接近于柏科。因此在分类学上的位置,曾被认为介乎杉科和柏科之间,而单列为水杉科(Metasequoiaceae)。
杉科植物,我国引种栽培的还有:巨杉属的巨杉(世界爷) [Sequoiadendron giganteum(Lindl.)Buchholz],常绿高大乔木,高达142米,胸径12米,树龄在3500年以上,原产北美。北美红杉属的北美红杉(长叶世界爷)[Sequoia sempervirens(Lamb.)Lindl.],常绿高大乔木,高可达104米,最高达110米,胸径8米,树龄达4000年以上,原产北美。巨杉、红杉都是北美单种属。
本科植物以种鳞和苞鳞半合生,种鳞具2—9粒种子,叶披针形、钻形、条形或鳞状,互生,螺旋状排列或2列(除水杉属对生外),小孢子无气囊等为特色。

(三)柏科(Cupressaceae)
常绿乔木或灌木。叶交互对生或轮生,稀螺旋状着生,鳞形或刺形,或同一树上兼有两型叶。孢子叶球单性,同株或异株。小孢子叶交互对生,小孢子囊常多于2个,小孢子无气囊。珠鳞交叉对生或3—4片轮生,珠鳞腹面基部有1至多枚直立胚珠,苞鳞与珠鳞完全合生。球果通常圆球形,种鳞盾形,木质或肉质,熟时张开或肉质合生呈浆果状。种子两侧具窄翅或无翅,或上端有1长1短的翅。染色体:X=11。
本科共22属,约150种,分布于南、北两半球。我国产8属,29种,分布几遍全国,另引入栽培1属,15种。
1.侧柏属(Platycladus) 生鳞叶的小枝扁平,排成一平面,直展或斜展。叶鳞形交叉对生。孢子叶球单性同株,单生于短枝顶端。大孢子叶球有4对交互对生的珠鳞,仅中间2对各生1—2枚胚珠。球果当年成熟,熟时裂开,种鳞木质、扁平,背部近顶端具反曲的钩状尖头,种子无翅,稀有极窄的翅。仅有侧柏[P.orientalis(Linn.)Franco]1种(图7-14,A),我国特产,除新疆、青海省外,分布几遍全国。材质细密,坚实,可供建筑等用材;枝叶药用,能收敛止血、利尿健胃;种子可榨油,入药滋补强壮、安神润肠;树姿优美,常栽培作庭园树种。
2.柏木属(Cupressus) 小枝斜上伸展,稀下垂,生鳞叶的小枝四棱形或圆柱形,不排成一平面,稀扁平而排成一平面。叶鳞形,交叉对生,排成4行,同型或2型,仅幼苗或萌生枝之叶为刺形。孢子叶球单性同株,单生枝顶。球果第二年成熟,种鳞4—8对,熟时张开,木质,盾形,顶端中部常具凸起的短尖头。种子具棱,两侧有窄翅。
本属约20种,分布于北美、东亚及地中海。我国产5种,分布于秦岭及长江流域以南各省,另引入栽培4种。柏木(C.funebris Endl.)(图7-14,B),生鳞叶小枝扁平,排成平面,下垂,球果小,直径0.8—1.2厘米,每种鳞具5—6枚种子。我国特有树种,分布很广,产于华东、中南、西南以及甘肃、陕西南部。材质优良,可供建筑、桥梁、造船、家具等用材;枝叶可提取芳香油;亦栽培作园林绿化及观赏树种。干香柏(C.duclouxiana Hickel),生鳞叶小枝四棱形,不下垂,球果大,直径1.6—3厘米。我国特有树种,产于云南中部及四川西部。
3.圆柏属(Sabina) 叶刺形或鳞形,或同一植株上兼有鳞形及刺形叶。孢子叶球单性异株或同株,单生于枝顶。球果熟时种鳞愈合,肉质,不张开。种子无翅。约50种,分布于北半球。我国产15种,另引入栽培2种。常见的有圆柏[S.chinensis(Linn.)Ant.](图7-14,C),叶兼有鳞叶与刺叶,刺叶3叶轮生,小枝不下垂。原产我国,分布于华北、华东、西南及西北等省区。朝鲜、日本也有分布。木材坚韧耐用,有香气,可供建筑等用材;枝叶及种子可提取挥发油和润滑油。
4.刺柏属(Juniperus) 冬芽显著。叶全为刺形,3叶轮生,基部有关节。大孢子叶球有3枚轮生的珠鳞,胚珠3枚,生于珠鳞之间。球果近球形,浆果状,熟时种鳞合生,肉质、不张开。种子无翅。有10余种,分布于亚洲、欧洲及北美。我国产3种,另引入栽培1种。常见有刺柏(J.formosana Hayate),我国特产,分布很广,木材可供建筑、家具等用,树形美观,各地均有栽培作庭园树种。
本科植物在我国常见的还有福建柏[Fokienia hodginsii(Dunn.)Henry et Thomas],为我国特产,分布于福建、浙江等省。日本花柏[Chamaecyparis pisifera(Sieb.et Zucc.)Endl.]和日本扁柏[C.obtusa(Sieb. et Zucc.) Endl.],均自日本引种,作观赏树。
本科植物以叶对生或轮生,具两型叶,种鳞和苞片完全合生,珠鳞交互对生或3—4片轮生,胚珠直立等为特色。
此外,分布于南半球热带及亚热带地区的南洋杉科(Araucariaceae),共2属,约40种。我国引入栽培2属,4种,如南洋杉(Araucaria cunninghamia Sweet)和异叶南洋杉[A.heterophylla(Salisb.)Franco],均原产大洋洲,我国南方常栽培作庭园树。
第五节 红豆杉纲(紫杉纲)(Taxopsida)
常绿乔木或灌木,多分枝。叶为条形、披针形、鳞形、钻形或退化成叶状枝。孢子叶球单性异株,稀同株。胚珠生于盘状或漏斗状的珠托上,或由囊状或杯状的套被所包围。种子具肉质的假种皮或外种皮。
在传统的分类中,本纲植物通常被放在松柏纲(目)中,但根据它们的大孢子叶特化为鳞片状的珠托或套被,不形成球果以及种子具肉质的假种皮或外种皮等特点,从松柏纲中分出而单列1纲。
红豆杉纲植物有14属,约162种,隶属于3科,即罗汉松科、三尖杉科和红豆杉科。我国有3科,7属,33种。这3科在系统发育上有紧密关系,可能来自共同的祖先。
(一)罗汉松科(Podocarpaceae)
常绿乔木或灌木。管胞具单列稀2列的缘孔,木质射线单列,无树脂道。叶常为条形、披针形,稀为鳞状钻形,或退化成叶状枝,螺旋状散生,稀近对生。孢子叶球单性异株,稀同株;小孢子叶球穗状,单生或簇生于叶腋,或生于枝顶,小孢子叶多数,螺旋状排列,各有2个小孢子囊,小孢子通常有气囊;大孢子叶球单生叶腋或苞腋,或生于枝顶,稀穗状,具多数至少数螺旋状着生的苞片,部分或全部或仅顶端之苞腑着生1枚倒生的胚珠(稀有为直生的),胚珠为1囊状或杯状的套被所包围,稀无套被。雄配子体的营养细胞6—8个或4—6个,精子2个,通常仅1个具有机能,另1个很快衰退。雌配子体有发达的大孢子囊,颈卵器2个,稀3—5个或更多。种子核果状或坚果状,成熟时,珠被分化成薄而骨质的外层和厚而肉质的内层2层种皮,套被变为革质的假种皮;或珠被变成极硬而骨质的种皮,套被变成肉质的假种皮,有时苞片与轴愈合发育成肉质种托。染色体:X=9-13、15、17、19。
本科共8属,约130种。分布于热带、亚热带及南温带地区,在南半球分布最多。我国产
1.罗汉松属(Podocarpus) 大孢子叶球生于叶腋或苞腋,套被与珠被合生,种子当年成熟,核果状,常有梗。全部为肉质假种皮所包,常生于肉质肥厚或微肥厚的种托上,稀苞片不发育成肉质种托。约100种,主要分布于南半球。我国有13种,3变种,分布于长江以南各省和台湾省。常见的有竹柏[P.nagi(Thunb.)Zoll.et Mor.]。叶对生,革质,卵形或卵状披针形,有多数并列的细脉,无中脉。小孢子叶球穗状圆柱形,单生叶腋,常呈分枝状。材质细致均匀,硬度中等,可供建筑、家具、文具、工艺等用材;种子榨油供食用或工业用。罗汉松[P.macrophyllus(Thunb.)D.Don](图7-15),叶条状披针形,中脉显著隆起。种子卵圆形,成熟时呈紫色,颇似一秃顶的头,而其下的肉质种托,膨大呈紫红色,仿佛罗汉袈裟,故名罗汉松。产于江苏、浙江、云南、广西等省,鸡毛松(P.imbricatus Bl.),叶小,异型,钻状条形叶排成两列,鳞状叶覆瓦状排列。种子顶生,种托红色、肉质。产于海南岛、广西及云南等地。
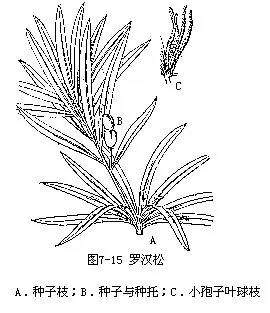
2.陆均松属(Dacrydium) 大孢子叶球生于小枝顶端,套被与珠被离生。种子坚果状,仅基部为杯状肉质或较薄而干的假种皮所包,苞片不增厚成肉质种托。约20种,主要分布南半球热带地区,我国仅有陆均松(D.pierrei Hickel)1种。产海南省。
(二)三尖杉科(粗榧科)(Cephalotaxaceae)
常绿乔木或灌木,髓心中部具树脂道。管胞具单列纹孔及2条或1条大型螺纹增厚。小枝近对生或轮生,基部有宿存芽鳞。叶条形或披针状条形,交互对生或近对生,在侧枝上基部扭转排成两列。孢子叶球单性异株,稀同株;小孢子叶球6-11聚生成头状,每个小孢子叶球由4—16个小孢子叶组成,各具2—4个(通常为3)小孢子囊,小孢子球形,无气囊;大孢子叶变态为囊状珠托,生于小枝基部(稀近枝顶)苞片的腋部,成对组成大孢子叶球,由3—4对交互对生的大孢子叶球组成大孢子叶球序。种子第二年成熟,核果状,全部包于由珠托发育成的肉质假种皮中,外种皮质硬,内种皮薄膜质,胚具子叶2枚。染色体:X=12。
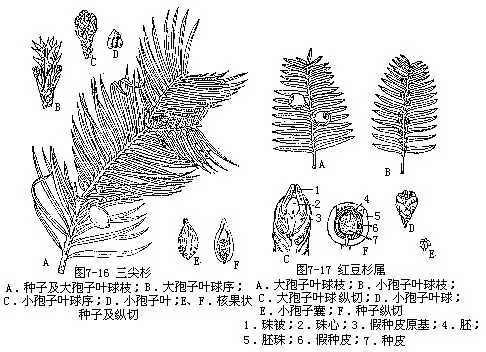
本科仅有三尖杉属(粗榧属)(Cephalotaxus)1属,9种。我国产7种,3变种。常见的有三尖杉(C.fortunei Hook.f.)(图 7-16),叶长 4—13厘米,宽3.5—4.4毫米,先端渐尖成长尖头。小孢子叶球有明显的总梗,长约6—8毫米。为我国特有树种,分布较广。材质优良,富有弹性,可供制农具、文具、工艺等用;枝、叶、根、种子可提取多种植物碱,供制抗癌药物;种子榨油供工业用;树冠优美,可作庭园树种。粗榧[C.sinensis(Rehd. et Wils.)Li],叶较短,长2—5厘米,宽约3毫米,先端常渐尖或微凸尖,基部近圆形,几无柄。小孢子叶球总梗长约3毫米。为我国特有树种,第三纪孑遗植物。分布很广。
三尖杉属在系统发育上与罗汉松属的原始代表密切相关,是罗汉松科与红豆杉科之间的一个中间环节。在北半球的白垩纪和第三纪地层中曾发现它们的代表。
(三)红豆杉科(紫杉科)(Taxaceae)
常绿乔木或灌木。管胞具大形螺纹增厚,木射线单列,无树脂道。叶条形或披针形,螺旋状排列或交互对生,叶腹面中脉凹陷,叶背沿凸起的中脉两侧各有1条气孔带。孢子叶球单性异株,稀同株;小孢子叶球单生叶腋或苞腋,或组成穗状花序状球序集生于枝顶,小孢子叶多枚,各有3—9个小孢子囊,小孢子球形,无气囊,外壁具颗粒状纹饰,单核状态时即传播;大孢子叶球通常单生,或少数2—3对组成球序,生于叶腋或苞腋,基部具多数覆瓦状或交互对生的苞片,胚珠1枚,基部具辐射对称的盘状或漏斗状珠托。雄配子体完全没有营养细胞。雌配子体具1—3或8个颈卵器。成熟种子核果状或坚果状,包于肉质而鲜艳的假种皮中。染色体:X=11、12。
本科有5属,约23种,主要分布于北半球。我国有4属,12种及1栽培种。
1.红豆杉属(紫杉属),(Taxus)(图7-17) 小枝不规则互生。叶条形,螺旋状着生,背面有两条淡黄色或淡灰绿色的气孔带,叶内无树脂道。孢子叶球单生叶腋。种子当年成熟,坚果状,生于杯状肉质的假种皮中,上部露出,成熟时肉质假种皮红色。约11种,分布于北半球。我国有4种,广布全国。红豆杉[T. chinensis(Pilger)Rehd.],为我国特有树种,第三纪孑遗植物,甘肃南部,陕西、四川、云南、湖南、湖北、广西、安徽等省均有。材质结构细致,防腐力强,为水上工程优良木材;种子含油60%以上,供制皂、润滑油及药用;叶常绿、深绿色、假种皮肉质红色,颇为美观,可作庭园树。
2.白豆杉属(Pseudotaxus) 本属与红豆杉属的主要区别是小枝近对生或近轮生。叶背面有两条白色气孔带,种子成熟时,肉质假种皮白色。仅有白豆杉[P.chienii(Cheng) Cheng]1种,为我国特有的单属种,产于浙江、湖南、广东、广西等省。木材供雕刻及器具等用。
3.穗花杉属(Amentotaxus) 叶交互对生,有树脂道。小孢子叶球多数,聚生成穗状花序状,常2—4穗生于近枝顶之苞腋;大孢子叶球单生于新枝上的苞腋或叶腋,有长梗。种子除顶端尖头裸露外,几全为囊状鲜红色肉质假种皮所包,基部有宿存的苞片。本属为我国特有属,共3种,分布于我国南部、中部、西部及台湾省南部。穗花杉[A.argotaenia(Hance)Pilger],叶下面白色气孔带通常与绿色边带等宽或较窄,穗状小孢子叶球通常2穗,长2—2.5厘米。为我国特有树种,产于江西、四川、西藏及华中、华南等地。木材结构细致,耐腐力强,供雕刻、模型、工艺品等用材;种子含油50%,可制肥皂等;叶常绿,较长,种子大,成熟时假种皮鲜红色,垂于绿叶之间,极为美观,可作庭园树种。云南穗花杉(A.yunnanensis Li),叶下面淡褐色或淡黄白色气孔带较绿色边带宽;小孢子叶球聚成穗状,通常4或4穗以上,长10—15厘米。为我国特有树种,产于云南东南部。台湾穗花杉(A.formosana Li),我国特有树种,产台湾省南部。
4.榧树属(Torreya) 枝轮生;小枝近对生或近轮生,基部无宿存芽鳞。叶交互对生或近对生,先端有刺状尖头;叶面中脉不明显,背面有两条较窄的气孔带;叶内有树脂道。小孢子叶球单生叶腋;大孢子叶球两个成对生于叶腋,胚珠1个,生于漏斗状的珠托上。种子第二年成熟,核果状,全部包于肉质假种皮中,基部有宿存的苞片。共7种,分布于我国、日本及北美。我国产4种,引入栽培1种。香榧(榧树)(T.grandis Fort.),叶先端有凸起的刺状短尖头,基部圆或微圆,长1.1—2.5厘米。为我国特有树种,产华东、湖南及贵州等地。材质优良,可作土木建筑及家具等用材;香榧的种子“香榧子”为著名的干果,亦可榨油,供食用;假种皮与叶,可提取香榧油,供药用。日本榧(T.nucifera Sieb. et Zucc.),叶先端有较长的刺状尖头,基部微圆或楔形,长2—3厘米。原产日本,我国各大城市有引种栽培,作庭园树。
第六节 买麻藤纲(倪藤纲)(Gnetopsida)
[盖子植物纲(Chlamydospermopsida)]
灌木或木质藤本,稀乔木或草本状小灌木。次生木质部常具导管,无树脂道。叶对生或轮生,叶片有各种类型;有细小膜质鞘状,或绿色扁平似双子叶植物;也有肉质而极长大,呈带状似单子叶植物。孢子叶球单性,异株或同株,或有两性的痕迹,孢子叶球有类似于花被的盖被,也称假花被,盖被膜质、革质或肉质;胚珠1枚,珠被1—2层,具珠孔管(micropylar tube);精子无纤毛;颈卵器极其退化或无;成熟大孢子叶球球果状、浆果状或细长穗状。种子包于由盖被发育而成的假种皮中,种皮1—2层,胚乳丰富。
买麻藤纲植物共有3目,3科,3属,约80种。我国有2目,2科,2属,19种,分布几遍全国。这类植物起源于新生代。茎内次生木质部有导管,孢子叶球有盖被,胚珠包裹于盖被内,许多种类有多核胚囊而无颈卵器,这些特征是裸子植物中最进化类群的性状。
(一)麻黄科(Ephedraceae)
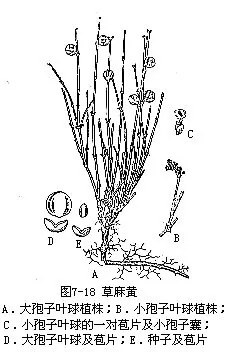
灌木、亚灌木或草本状,多分枝,小枝对生或轮生,具节。节间有多条细纵槽纹,横切面常有棕红色髓心。叶退化成鳞片状,对生或轮生,2—3片合生成鞘状,先端具三角状裂齿。孢子叶球单性,异株稀同株;小孢子叶球单生,或数个丛生;或3—5个成复穗状,具膜质苞片数对,每苞片生一小孢子叶球,其基部具2片膜质盖被及一细长的柄,柄端着生2—8个小孢子囊,小孢子椭圆形,具5—10条纵沟槽;大孢子叶球有数对交互对生或3片轮生的苞片,仅顶端1—3苞片生有1—3枚胚珠,每个胚珠均由1层较厚的囊状盖被包围着,胚珠具1—2层膜质珠被,珠被上部(2层者仅内被)延长成充满液体的珠孔管。小孢子萌发基本上与松属相似,最后形成1个管细胞核和1个生殖细胞核,后者再分裂为1个足核和1个精核,精核分裂成2个无鞭毛的精子。成熟的雌配子体充满了细胞,通常有2个(有时1个或3个)颈卵器,颈卵器具有32个或更多的细胞构成的长颈,中央细胞核分裂为卵核和腹沟细胞核。种子成熟时,盖被发育为革质或稀为肉质的假种皮,大孢子叶球的苞片,有的变为肉质,呈红色、桔红色或橙黄色,包于其外面,呈浆果状,俗称“麻黄果”;有的则变为干膜质甚至木质化。染色体:X=7。
本科隶属于麻黄目(Ephedrales),仅麻黄属(Ephedra)1属,约40种,分布于亚洲、美洲、欧洲东南部及非洲北部干旱、荒漠地区。我国有12种及4变种,分布较广,以西北各省区及云南、四川、内蒙等地种类较多。本属植物,多数种类含有生物碱,为重要的药用植物;生于荒漠及土壤瘠薄处,有固沙保土作用;“麻黄果”供食用。草麻黄(E.sinica Stapf. )(图 7-18),植株无直立木质茎,呈草本状,小枝节间较长。大孢子叶球成熟时矩圆状卵圆形或近圆球形,种子常2粒。广布于我国东北、华北及西北等省区,习见于山坡、平原、干燥荒地及草原等处。木贼麻黄(E.equisetina Bunge),有直立木质茎,呈灌木状,节间细而较短,小孢子叶球有苞片3—4对;大孢子叶球成熟时长卵圆形或卵圆形。种子通常1粒。产于内蒙、河北、山西、陕西、甘肃及新疆等地,习见于干旱地区的山脊山顶或石壁等处。
(二)买麻藤科(Gnetaceae)
大多数为常绿木质大藤本,极少为直立灌木或乔木,茎节由上、下两部接合而成,呈膨大关节状。次生木质部具多列圆形具缘纹孔的管胞和导管,宽的射线,厚的皮部及多数粘液沟,以及位于韧皮部外侧的针状细胞层。单叶对生,椭圆形或卵形,革质或半革质,具羽状侧脉及网状细脉,极似双子叶植物。孢子叶球单性,异株,稀同株;孢子叶球序伸长成细长穗状,具多轮总苞,总苞浅杯状,由多数苞鳞愈合而成。小孢子叶球序单生或数个组成顶生或腋生聚伞花序状,各轮总苞有多数小孢子叶球,排成2—4轮,小孢子叶球具管状盖被,每个小孢子叶,1—2个或4个小孢子囊,小孢子圆形。大孢子叶球序每轮总苞内有4—12个大孢子叶球,各具2层盖被,外盖被极厚,是由2个盖被片合生而成,内盖被是外珠被,珠被的顶端延长成珠孔管,大孢子囊内通常有2—3个大孢子母细胞,各自形成4个大孢子,其中若干个能够发育为雌配子体,但除了最下面1个雌配子体继续发育外,其他的均退化;雌配子体在受精前始终处于自由核状态;到受精时分化出来1、2或3个游离核,在其周围形成细胞质,而近似于被子植物的卵核,不形成颈卵器,这是裸子植物(颈卵器植物中发展到最高水平的一群)中的例外情况,可称之为“没有颈卵器的颈卵器植物”。小孢子萌发成4核状态时,似乎是由昆虫传至珠孔管分泌的传粉滴上,随着滴液的干涸而被吸入珠孔管中,并形成花粉管,花粉管生长到达雌配子体时,2个精子,管核和一些细胞质即流入雌配子体,2个精子向卵核移动,其中1个与卵核结合。胚的发育无游离核阶段,具有发达的胚足,长的胚轴和2枚子叶。在配子体中,虽可有数个卵核同时受精,但最后只有1个胚发育成熟。种子核果状,包于红色或桔红色肉质假种皮中,胚乳丰富。染色体:X=11。
本科隶属于买麻藤目(Gnetales),仅买麻藤属(Gnetum)(图7-19)1属,共30余种,分布于亚洲、非洲及南美洲等的热带及亚热带地区。我国有1属,7种,分布于福建、广西、贵州、云南、江西及湖南等省区,最北分布在福建约达北纬26.6°处,这也是现在全世界已知的最北记录。买麻藤属植物茎皮富含纤维,为织麻袋,鱼网、绳索等原料;种子可炒食或榨油,供食用或作机器润滑油。常见的有买麻藤(G.montanumMarkgr.),大藤本,高达10米以上。叶通常呈矩圆形,革质或半革质,长10—25厘米,宽4—11厘米。小孢子叶球序1—2回三出分枝,排列疏松。成熟种子,常有明显种子柄。小叶买麻藤[G.parvifolium(Warb.)C.Y.Cheng],缠绕藤本,高达4—12米,叶椭圆形,革质,长4—10厘米,宽2.5厘米。小孢子叶球序不分枝或一次分枝。成熟种子无种柄或近无种柄。

(三)百岁兰科(Welwitschiaceae)
植物体的形态非常奇特(图7-20,21),不同于其他裸子植物。茎部粗短,块状,有圆锥根深入地下。植物体除了在幼苗时期还有1对子叶(2—3年后脱落)外,终生只有1对大型带状叶子,长达2—3米,宽约30厘米,可生存百年以上,故名百岁兰。孢子叶球序单性异株,生于茎顶凹陷处;孢子叶球序的苞片交互对生,排列整齐,呈鲜红色。小孢子叶球具6个基部合生的小孢子叶,中央有1个不完全发育的胚珠。这说明百岁兰是来自两性花的祖先。大孢子叶球盖被筒状,胚珠具一层珠被,珠被顶端延伸成珠孔管。百岁兰也没有颈卵器,其受精作用是由雌配子体的顶部向上生出的管状突起和珠心中向下生长的花粉管相遇而进行的。染色体:X=21。
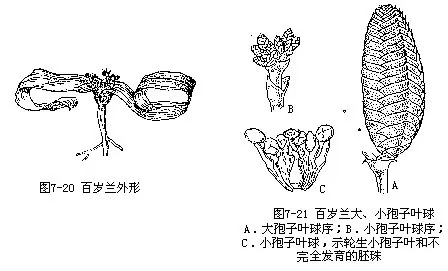
本科隶属于百岁兰目(Welwitschiales),仅百岁兰属(Welwitschia)1属,百岁兰[W.bain-esii(Hk.f.)Carr.](图7-20,21)1种,为典型的旱生植物,分布于非洲西南部,靠近海岸的沙漠地带。
第七节 裸子植物的起源与进化
裸子植物在系统发育过程中,植物体的次生生长由微弱到强;茎干由不分枝到多分枝;孢子叶由散生到聚生成各式孢子叶球;大孢子叶逐渐特化;雄配子体由吸器发展为花粉管;雄配子由游动的、多纤毛精子,发展到无纤毛的精核;颈卵器由退化、简化发展到没有等等。这一系列的发展变化都是和系统演化密切相关。尤其是生殖器官的演化,使裸子植物有可能更完善地适应陆生生活条件,而达到较高的系统发育水平。
在二叠纪的早期,亚洲、欧洲和北美部分地区开始出现酷热、干旱的气候环境,许多在石炭纪繁盛一时的造煤植物,因不能适应自然环境的变化,而趋于衰落和绝灭。而一群以种子繁殖的高等植物,即裸子植物,因适应当时自然环境的变化却得到了发展而繁荣兴旺,成为当时地球上植被的主角。
古生代的石炭纪、二叠纪是地球上蕨类植物、种子蕨和苛得狄植物(Cordaitinae)繁荣昌盛时期。随着岁月的流逝,自石炭纪的中、晚期起,地球上由于气候和其他自然因素的影响,丛林中的面貌,即植被也在发生变化,逐渐形成了4个不同的植物群:分布在欧洲、北美洲大部地区的称为欧美植物群;发育在亚洲东部的就称为华夏植物群[大羽蕨(Gigantopteris)]。欧美植物群和华夏植物群生长于气候湿热的条件,植被与今日的雨林、季雨林相似。在亚洲北部季节明显、湿度高而温度较低的生境,分布着安加拉植物群(或称通古斯植物群、库兹涅茨克植物群)和在南半球各大洲和北半球南亚地区季节明显,湿度和温度变化显著的环境,分布着冈瓦纳植物群[舌蕨(Glossopteris)]。
在石炭纪和二叠纪之交,地球上自然环境开始发生了一系列的变化,华夏植物群和欧美植物群分布的地区先后出现了季节性的干旱,并逐渐增加着强度和幅度,严重地威胁着生长在湿润环境中的各种植物。与此同时,大规模的地壳运动,使陆地上升,面积和相对高度迅速增加,大片的沼泽干涸或消失。又随着海水的退却,滨海湿润而均匀的海洋性气候,也被严酷而多变的大陆性气候所代替,这些自然因素的变化,对于植物界的影响,更起了推波助澜的作用。盛极一时的蕨类植物大量衰亡,新型的裸子植物逐渐兴旺起来。
裸子植物虽然到古生代末期之后,方始形成为陆地植物中的主要代表,它的历史可远溯到3.5亿年前,也就是地质史上称为中、晚泥盆世的时候。从化石记载表明,那时裸子植物正处于形成和开始发展阶段。最古老的裸子植物或称原裸子植物(Progymnosperm),因为它们尽管在某些方面比蕨类植物进化,但尚未具备裸子植物全部的基本特征。下面我们从在地层中保存下来的生物历史的化石记录,简述裸子植物的发生和发展(图7-22)。

无脉蕨(Aneurophyton)是中泥盆世的一种原裸子植物,树干高、茎粗的乔木,茎顶端有1个由许多分枝组成的树冠,它的末级“细枝”形状就像分叉的叶片,但其中无叶脉。孢子囊小而呈卵形,生于末级“细枝”之上。茎干内部具次生木质组织,这种组织由具缘纹孔的管胞组成。它没有发达的主根,只有许多细弱的侧根。
古蕨(Archaeopteris)是晚泥盆世特有的一群较为进化的原裸子植物的代表。树高、茎粗的塔形乔木,茎干具有次生生长的组织,输导组织中的木质成分是具缘纹孔的管胞,茎干的顶端有1个由枝叶组成的树冠;叶是扁平而宽大的羽状复叶;根系较无脉蕨发达;孢子囊单个或成束地着生在不具叶片的小羽片上,孢子囊内曾发现大、小两种孢子。
无脉蕨、古蕨这一类十分奇特的植物,却具有大、小孢子,羽状复叶,具缘纹孔的管胞等原裸子植物的重要特征。所以,被认为与裸子植物的祖先有关。但是它们没有胚珠更没有种子,大概是原始蕨类向着原裸子植物演进的低级的过渡类型。1974年伯恩(Burn)将古蕨算作原裸子植物。到了石炭纪、二叠纪时,从原裸子植物繁衍出更高级的类型。
裸子植物的进一步繁演就是种子蕨(Pteridospermae)的发现。种子蕨在上泥盆世地层中已发现,上石炭纪是其极盛时期,少数代表曾生存到三叠纪末期之前。在1903年英国的古植物学家发现了凤尾松蕨(Lyginopteris oldhamia)的“真蕨”,竟然以种子来进行繁殖,是当今知道得最清楚的种子蕨(图7-23)。叶为多回羽状复叶,甚大,叶轴上部分二叉;茎甚细,髓颇大,有形成层,维管束的大部分由次生木质部和次生韧皮部组成;小型的种子,外有1杯状包被,其上生有腺体,种子中央为1颇大的雌配子体组织和颈卵器,珠心的顶端有1突出的喙,喙外又有一垣围之,二者之间为贮粉室,其中有时可看见花粉粒,珠心之外有1厚的珠被,珠被是由若干个单位联合而成的,每一个单位代表着1个不育的大孢子叶,所以整个胚珠不是单个的孢子叶,而是聚合囊(synangium),珠心才是有效的大孢子囊。

在石炭纪、二叠纪化石中,还发现以髓木(Medullosa)为代表的植物,也是当时北半球广泛分布的种子蕨。在我国地质史的石炭纪、二叠纪时期,也有许多种子蕨繁盛生长,不仅有凤尾松蕨和髓木的家族成员,还有若干特殊的类型。最著名的是大羽蕨(Gigantopteris),这种植物的整个形状和内部结构虽然不很清楚,但从叶的形态特征来看,很可能是一种比较进化的种子蕨,它的叶子像被子植物茄科的烟草叶,长可达数十厘米,叶脉也相似,都是属于“复杂网状脉序”。大羽蕨是迄今所知具有这种高级脉序的先驱者。由于我国和东亚地区在二叠纪时,繁荣着以大羽蕨为代表的独特的植物群,故称之为华夏植物群。
在这一类群最古老、最原始的裸子植物中,有几个方面值得特别注意:(1) 它们还没有花,但已形成种子,这在植物系统发育过程中,种子的出现比花和果实更早;(2) 在种子中始终没有发现发育完善的胚,这是一种原始的性状;(3) 在胚珠的贮粉室中,只有看到花粉粒,而未发现花粉管,这也是原始的性状之一。所以,种子蕨是介于蕨类植物和裸子植物之间的1个极其重要的类型,并成为许多现代裸子植物的起点。
拟铁树植物(Cycadeoideinae)即本内铁树(Bennettitinae)和科得狄植物的发现,对由种子蕨植物直接演化出来的铁树植物和一些古植物,具有重要意义。
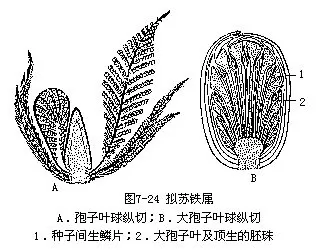
拟苏铁植物,是直接起源于髓木类种子蕨植物。其中拟铁树属(Cycadeoidea)(图7-24),植物体皆矮小,茎有块状或柱状,茎的解剖构造上有1大髓,1薄层维管组织及1厚的皮层,茎的表面覆满已脱落的叶基,茎顶端有1丛羽状复叶。孢子叶球遍布于茎的周围;孢子叶球两性,即大、小孢子叶合成 1孢子叶球;在孢子叶的下部为螺旋状排列的苞片,其上为1轮大型羽状的小孢子叶,基部相连成盘,小孢子囊排成两列,每个小孢子囊又分数室,为聚合囊;每个大孢子叶只有1个短柄和1个顶生的胚珠夹于大孢子叶之间,又有另1种苞片,棒形,顶端膨大,称为间生鳞片(interseminal scale)。种子无胚乳,而含有2片子叶的胚。拟铁树属植物,极似铁树植物,但孢子叶球两性,成熟种子无胚乳,这在裸子植物中颇为特殊。因此,被认为是和某些具有两性结构的裸子植物的起源有关的一群古植物。
科得狄植物,植物体为高大乔木,茎粗一般不超过1米,茎干的内部构造和种子蕨颇相似。但木材较发达而致密,木质部或薄或厚,通常无年轮,构造特殊的髓心,是由许多薄壁细胞形成的横裂成片,仿佛似被子植物胡桃的髓;具较发达的根系和高大的树冠;叶皆是全缘的单叶,形态大、小颇不一致,其上有许多粗细相等、分叉的、几乎是平行的叶脉,脉间尚有硬组织形成细纹;已有了“花”,即孢子叶球,大、小孢子叶球分别组成松散的孢子叶球序。小孢子叶球的基部有多数不育的苞片,小孢子叶由小孢子叶柄和小孢子囊组成。大孢子叶的结构与小孢子叶相似,基部具不育的苞片,胚珠顶生,珠心和珠被完全分离。有胚珠,但还没有真正的种子,有贮粉室,其内曾多次发现有花粉粒,但未发现有花粉管(图7-25)。

科得狄植物具胚珠,叶的形态、结构等类似种子蕨。而茎的构造和孢子叶球等又类似裸子植物。它是种于蕨的后裔或具有共同的起源。它在裸子植物的起源和系统发育上都具有重要的意义。
关于真蕨类(Filicinae)、原裸子植物和裸子植物的系统发育,60年代贝克(Beck)把古蕨属和科得狄属的植物归属原裸子植物,并认为可能是种子植物的直接祖先;70年代班克(Bank)提出真蕨、原裸子植物都是来自裸蕨(Psilophyton),再由原裸子植物进化到裸子植物(图7-26)。

综上所述,现代裸子植物的起源和进化,根据地质年代的历史记载和古植物学的研究,概述于下。
铁树纲植物 铁树纲植物起源开始于古生代二叠纪,甚至可能起源于石炭纪,繁盛于中生代,是现代裸子植物最原始的类群。从种子蕨的发现、研究表明,它们有着密切的关系。在形态上,茎干都不甚高大,少分枝或不分枝,茎干表面残留叶基,顶生一丛羽状复叶;内部构造上,都具有较大的髓心和厚的皮层,木材较疏松;生殖器官结构上,小孢子叶保存着羽状分裂的特征,大孢子叶的两侧着生数个种子,呈羽状排列;它们的种子结构也很接近。这些都说明铁树类植物是由种子蕨演化而来的。
银杏纲植物 地质历史时期植物化石的研究,提供了可靠而丰富的依据。从化石材料记载,它的历史可远溯至石炭纪,晚石炭纪出现的二歧叶(Dichophyllum)。之后,早二叠纪的毛状叶(Trichopitys),晚二叠纪的拟银杏(Ginkgoites)、拜拉(Baiera),三叠纪的楔银杏(Sphenobaiera)等或许是银杏的远祖。到了中侏罗世已有许多银杏生存。又从楔银杏、拜拉的孢子叶的情况看,它们的小孢子叶上有5—6个(偶有3—7个)小孢子囊,而银杏有3个小孢子囊;毛状叶、拜拉和拟银杏等的大孢子叶上的胚珠数目,也多于现代的银杏。由此看来,现存银杏的小孢子囊和大孢子囊,可能是经历了一系列“简化”过程演变而成的。另一方面,银杏类和科得狄也有一些相似之处,比较重要的是它们单叶的叶基构造和叶脉形式一致;又科得狄的胚珠具有贮粉室,可能以游动精子进行受精等特点和银杏相似,这些都说明它们起源于共同的祖先。
红豆杉纲植物 从古植物学的研究,为我们提供了地质历史时期这类植物盛衰的情况和演化趋向的资料,但是由于化石材料的不完整和研究程度所限,现存的红豆杉纲各科、属和已灭绝的类型之间的演化线索,还未能完全搞清。一般认为红豆杉纲3个科:罗汉松科、三尖杉科(粗榧科)和红豆杉科(紫杉科),在系统发育上有密切关系。三尖杉科植物的孢子叶球中,没有营养鳞片,很可能是晚古生代的安奈杉(Ernestiodendron),通过中生代早期的巴列杉(Palissya)、穗果杉(Stachyotaxus)的途径演化而来的。而罗汉松科、紫杉科,则与科得狄植物有相似之处,尤其是大孢子叶球的结构以及变态的大孢子叶;穗状花序式的小孢子叶球序,保持着和科得狄类似的原始性状。说明这两个科的植物,可能是从科得狄类直接演化出来的。
松柏纲植物 松柏纲植物是现代裸子植物中种、属最多的植物。它们的植物体的形态结构比铁树类、银杏类更能适应寒旱的自然环境;它们的胚珠受精方式比较进化,小孢子(花粉粒)萌发时产生花粉管,游动精子消失。这是由于此类植物在地质历史进程中较能抵御自然环境的变动,而较多地保存下来的缘故。关于松柏类植物的起源,还不很清楚,在地质史上出现较早的科得狄,可看作是松柏类植物的先驱者,因为它和古老的松柏类在形态上和结构上,有不少重要的相似点,特别是和石炭纪、二叠纪的松柏植物勒巴杉(Lebachia)孢子叶球的结构非常近似。
从这些相似的特征分析,松柏纲植物,无疑地是科得狄的后裔。一般认为,松柏纲植物各科的演化路线是:杉科和柏科,它们可能是从中生代早期的三叠纪、侏罗纪时已灭绝的类型,伏脂杉(Voltzia)、掌鳞杉(Cheirolepis=Hirmerella)等化石类型中分化出来的;南洋杉科,在木材的形态结构上与科得狄极为相似,称为“南洋杉型”,所以,可能是从它直接演化出来的;松科的可靠化石,虽出现较晚,但也许很早就已形成为一个独立的演化支,因为它的球果具有分离的苞鳞,是相当原始的性状。
买麻藤纲植物 买麻藤纲植物在现代裸子植物中,是完全孤立的一群。现存的3个属即麻黄属、买麻藤属和百岁兰属,这3个属缺乏密切关系的类群,各自形成3个独立的科和目。它们在外形上和生活环境相差很大,地理分布上又较遥远。但从这3个属植物中,都可以或多或少地看到由生殖器官两性到单性,雌雄同株到异株的发展趋势,它们都是属于比较退化和特化的类型。
目前,对这个纲植物的历史还不很明了,根据它们形体的结构和明显的分节,被认为与木贼类植物有一定的亲缘关系;根据它们的孢子叶球的结构来看,其祖先曾具有两性的孢子叶球,而具有两性孢子叶球的,只有起源于种子蕨类的拟铁树植物,它们孢子叶球序二叉分枝,珠孔管、胚珠的特性等,说明买麻藤植物很可能是强烈退化和特化了的拟铁树植物的后裔;但是这个纲的植物,又都具有导管、精子无纤毛、颈卵器趋于消失,甚至受精作用是在雌配子体的自由核状态下进行的,这些特征又是堪与被子植物相比拟的高级性状。
裸子植物在其漫长的历史过程中,地史、气候经过多次的重大变化,裸子植物的种系也随之多次演替更新,老的种系相继绝灭,新的种系陆续演化出来,并沿着不同的进化路线不断地更新、发展、繁衍至今。
复习思考题
1.裸子植物有什么主要特征?它与苔藓植物和蕨类植物有什么共同点?有什么区别?
2.试以松属为例,简述松柏纲植物的生活史?
3.银杏、水杉均为我国特产,它们的发现在生物学上有什么重要意义?
4.试比较松科、杉科和柏科的异同点是什么?
5.为什么说买麻藤纲是裸子植物中最进化的类群?
6.简述裸子植物的起源与进化?
7.试述裸子植物在自然界中的作用及其在经济上的重要性。