一、花的概念和花的组成
(一)花的概念
种子植物从种子萌发时起,就不断地进行着生长和发育。在以营养生长为基础的前提下,经过一定的时期,满足了光照、温度等因素的要求,以及某些激素的诱导作用以后,就进入生殖生长阶段。这时一部分或全部茎端的分生组织,不再形成叶原基和腋芽原基,而是形成花原基或花序原基。
种子植物由营养生长转入生殖生长,是个体发育中的一个巨大转变,这一转变包含着一系列极为复杂的生理生化过程。转变开始时,茎端生长锥的表面有一层或数层细胞加速分裂,生长锥中部的细胞分裂慢,细胞变大,还出现大液泡。在表层细胞里没有淀粉的积累,但蛋白质和RNA含量很高,证明花的分化与DNA-RNA-蛋白质系列的活化有很大关系。此后,表层细胞由外而内地形成数轮突起,再由这些突起分别发展为组成花的各部分原基。
第一个为花下定义的人是德国的博物学家和哲学家哥德(J.W.von Goethe,1749—1832),在18世纪的90年代里,他提出植物一切器官的共同性的观点和多种多样植物形态的统一性的观点。按照他的观点,植物地上器官是统一的,是一种器官的多方面变态,于是,他提出花是适合于繁殖作用的变态枝。哥德的见解被认为基本上是正确的,因为花的各部分从形态、结构来看,还具有叶的一般性质。如果从高等植物系统进化的发展角度来看,作为产生大、小孢子的花的各部分,应该是由植物茎轴的孢子囊和孢子囊柄演变而来。
由此可以说,花是不分枝的变态短枝,用以形成有性生殖过程中的大、小孢子和雌雄配子的,并且进一步发展为种子和果实。所以花也是果实和种子的先导,花、果实、种子三者成为一体,但出现的先后和发展的性质以及结构互有不同。
种子植物的花可以有多方面的经济利用。由于花的鲜艳色彩和芬芳香味,利用花来美化环境,陶冶心情,已是尽人皆知的了。从花朵中提取芳香油料,制成香精,很早就受到重视,虽然有的香精可以人工合成,但一部分名贵的香料,仍然是从花朵中提制的。利用花朵如茉莉、代代、白兰花等熏制香茶,由来很早,已成为花茶制作过程中不可缺少的重要原料,有的花农专门栽植这类花卉植物,供制作香茶的需要。花朵用于医药方面的种类也很不少,常见的如红花、丁香、金银花、菊花等,都有较高的药用价值。少数植物的花朵可供作染料,如凤仙。有些植物的花朵或花序具有较高的营养成分,如金针菜、花椰菜等,或浓郁的香味,如桂花、玫瑰花等,可供食用或制作糕点。
(二)花的组成
一朵完整的花可分为五个部分,即花柄(pedicel)、花托(receptacle)、花被(perianth)、雄蕊群(androecium)和雌蕊群(gynoecium)(图4-12)。
1.花柄和花托 花柄或称花梗,是着生花的小枝,可以把花展布在枝条的显著位置上;同时,也是花朵和茎相连的短柄。花柄有长、有短,视不同植物种类而异,例如垂丝海棠的花柄很长,而贴梗海棠的花柄就很短,有些植物的花没有花柄。花柄有具分枝的,也有不分枝的。分枝的花柄称小梗,顶端着生一花。花柄的结构和茎的结构是相同的。
花托是花柄或小梗的顶端部分,一般略呈膨大状,花的其他各部分按一定的方式排列在它上面。花托的形状随植物种类而异,有的圆柱状,如木兰;有的凸起如覆碗状,如草莓;也有中央部分凹陷呈碗状,如桃、蔷薇等;或膨大呈倒圆锥形,如莲。柑橘的花托在雌蕊(pistil)基部形成膨大的盘状,能分泌蜜汁,称为花盘。此外,有的花托在雌蕊群基部向上延伸成为柄状,称雌蕊柄,如落花生,它的雌蕊柄在花完成受精作用后迅速延伸,将先端的子房插入土中,形成果实,所以也称为子房柄;西番莲、苹婆属等植物的花托,在花冠以内的部分延伸成柄,称为雌雄蕊柄或两蕊柄;也有花托在花萼以内的部分伸长成花冠柄,如剪秋萝和某些石竹科植物(图4-13)。
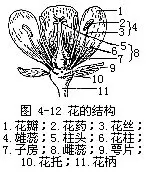

2.花被 花被着生在花托的外围或边缘部分,是花萼(calyx)和花冠(corolla)的总称,由扁平状瓣片组成,在花中主要是起保护作用,有些花的花被还有助于花粉传送。花被由于形态和作用的不同,可分为内、外二部分,在外的称花萼,在内的为花冠,像这样的花称两被花,如油菜、豌豆、番茄等。仅一轮花被的花称单被花如大麻、荞麦等。也有花被虽有二轮,但内、外瓣片在色泽等方面并无区分的,称为同被花,它们的每一瓣片称被片(tepal),如百合、丝兰等。也有花被完全不存在的,称为无被花,如杨、柳等。
(1)花萼 花萼是由若干萼片(sepal)组成,包被在花的最外层。普通萼片多为绿色的叶状体,在结构上类似叶,有丰富的绿色薄壁细胞,但无栅栏、海绵组织的分化。有的植物花萼大而具色彩,呈花瓣状,有利于昆虫的传粉,如飞燕草。棉的花朵除花萼外,外面还有一轮绿色的瓣片,称副萼(accessory calyx)。
萼片有各自分离的,称离生萼,如油菜、桃;也有彼此联合在一起成筒状,称为合生萼,如樱花。萼片大小相同或不同,分别称为整齐萼或不整齐萼。萼片比花冠先脱落的称早落萼,如罂粟;有的与花冠一起脱落,称落萼,如油菜、桃;也有花萼常留花柄上,随同果实一起发育,称宿萼,如茄、柿、番茄、辣椒等;有的在花萼一边引伸成短小的管状突起,称为距(spur),如凤仙花、旱金莲等植物的花萼。
(2)花冠 位于花萼的上方或内方,是由若干称为花瓣(petal)的瓣片组成,排列为一轮或多轮,结构上由薄壁细胞所组成(图4-14)。花瓣比萼片要薄,且多具鲜艳色彩。有的花瓣,表皮细胞呈凹凸不平状,经光线折射后,呈现丝绒般光泽。花瓣的色彩主要是因为花瓣细胞内含有色素所致。含杂色体时,花瓣呈黄色、橙色或橙红色;含花青素的花瓣显示红、蓝、紫等色(主要由液泡内细胞液的酸碱度所决定);有的花瓣两种情况都有存在,它的色彩往往就绚丽多彩。如果两种情况都不存在,花瓣便呈白色。花瓣基部常有分泌蜜汁的腺体存在,可以分泌蜜汁和香味。多种植物的花瓣细胞还能分泌挥发油类,产生特殊的香味。所以花冠除具保护雌、雄蕊的作用外,它的色泽、芳香以及蜜腺分泌的蜜汁,都有招致昆虫传送花粉的作用,为进一步完成有性生殖创造了有利条件。与萼片的情况相同,花瓣也有各自分离或互相联合的,具有分离花瓣的花称离瓣花,如桃、毛茛;具有联合花瓣的花为合瓣花,如牵牛、茄等。花瓣形状、大小相同的为整齐花,如桃;相反则为不整齐花,如豌豆。根据花瓣的脱落情况,有早落冠,如葡萄;落冠如桃和宿冠如黄瓜。花瓣有距的,如三色堇、耧斗菜等。

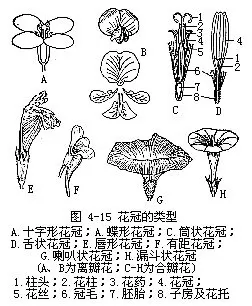
由于组成花冠的花瓣形状、大小相同或各异,花瓣各自分离或彼此联合,而用花冠形成多种不同的形状,有成十字形的,如油菜;有蝶形的,如豌豆;有漏斗状的,如牵牛;有钟状的,如风铃草;有管状的,如马兜铃;有唇形的,如野芝麻;有舌状的,如菊花等(图4-15)。
在蕾期,花冠的各瓣片因植物种类的不同而形成各种形式的卷叠。
3.雄蕊群 雄蕊群是一朵花中雄蕊的总称,由多数或一定数目的雄蕊(stamen)所组成,位于花被的内方或上方,在花托上呈螺旋或轮状排列。一般直接生于花托上,也有基部着生于花冠或花被上的。
少数原始被子植物的雄蕊呈薄片状(图4-16),可为花的雄蕊是叶的变态提供佐证,但绝大多数被子植物的雄蕊是由花丝和花药两部分组成(参阅图4-12)。花丝细长,顶端与花药相连。花药是产生花粉粒的地方,是雄蕊的主要部分,在结构上,由4个或2个花粉囊组成,分为两半,中间为药隔相连。
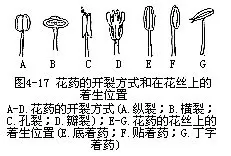
花粉成熟后,花粉囊自行破裂,花粉由裂口处散出。花粉囊破裂的方式,有的沿二粉囊交界处成纵行裂开,称为纵裂,如油菜、牵牛、百合等;有的沿花药中部成横向裂开,称横裂,如木槿、蜀葵等;有的在花药顶端开一小孔,花粉由小孔散出,称孔裂,如茄、番茄等;或在花药的侧壁上裂成几个小瓣,花粉由瓣下的小孔散出,称瓣裂,如香樟等(图4-17)。
花丝除一般细长如丝外,也有扁平如带的,如莲,或完全消失的,如栀子,或转化为花瓣的,如美人蕉。
花药在花丝上的着生方式也有几种不同情况:有的花丝顶端直接与花药基部相连,称为底着药(innate anther);有的花药背部全部贴着在花丝上的,称为贴着药(adnate anther);有的花丝顶端与花药背面的一点相连,整个雄蕊犹如丁字形的,称为丁字着药(versatile anther)。此外,如果花药向着雌蕊一面生长的称内向药(introrse),向着花冠生长的则称外向药(extrorse)。

雄蕊的数目和长短,随植物种类而异。原始种类植物的雄蕊数多而不一定,较进化的则有减少,以至达到一定的数目,此外,一朵花中的雄蕊一般长短相等,但也有同一花中的雄蕊长短不等,如十字花科植物的雄蕊共有6枚,其中外轮的2个较短,内轮的4个较长,称四强雄蕊(tetradynamous stamen);又如唇形科、玄参科植物的花朵中,雄蕊共有4枚,2个较长,另2个较短,称为二强雄蕊(didynamous stamen)。
雄蕊同样有分离和联合的变化,联合有在花丝部分,或在花药部分。花药完全分离而花丝联合成1束的称单体雄蕊(monodelphous stamen),如棉、山茶;花丝如联合成2束的称二体雄蕊(diadelphous stamen),如蚕豆、豌豆等;花丝合成为3束的为三体雄蕊(tridelphous sta-men),如小连翘;合为4束以上的为多体雄蕊(polydelphous stamen),如金丝桃。雄蕊的花丝分离而花药互相联合的称聚药雄蕊(synantherous stamen),如菊科、葫芦科等植物的雄蕊(图4-18)。


4.雌蕊群 雌蕊群是一朵花中雌蕊的总称,位于花中央或花托顶部。每一雌蕊由柱头(stigma)、花柱(style)和子房(ovary)三部分组成(参阅图4-12)。构成雌蕊的单位称为心皮(carpel),是具生殖作用的变态叶。有些植物,一朵花中的雌蕊只是由一个心皮所构成,称为单雌蕊(simple pistil),如蚕豆、大豆。更多种类植物的雌蕊是由几个心皮构成的,其中有的各心皮各自分离。因而各雌蕊也彼此分离,形成一朵花内多数雌蕊,称为离生雌蕊(apocarpous pistil),亦属单雌蕊,如玉兰、莲等;或是各个心皮互相联合,组成一个雌蕊,称为合生雌蕊(syncarpous pistil),属复雌蕊(compound pistil),以与单雌蕊相对应,如棉、番茄等。合生雌蕊各部分的联合情况不同,有子房、柱头和花柱全部联合的,也有子房和花柱联合而柱头分离的,也有只是子房联合而柱头、花柱彼此分离的(图4-19)。
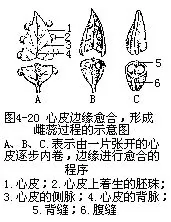
单雌蕊是通过心皮两侧边缘,向内包卷愈合而成(图4-20)。边缘愈合的部分形成一条缝线,称为腹缝,相对于心皮的中肋处的一条缝线,称背缝,在这二条缝线处都有维管束分布。在心皮联合的复雌蕊上,同样可以见到这两种缝线,但复雌蕊的腹缝位于2心皮彼此愈合的部分,如果复雌蕊是由3个心皮合成的,则可见到3条腹缝,相应的另有3条背缝,出现在3个心皮的背面中肋处。
(1)柱头 柱头位于雌蕊的顶端,是接受花粉的部位,一般膨大或扩展成各种形状。柱头的表皮细胞有延伸成乳头、短毛、或长形分枝毛茸的。当传粉时,有的柱头表面湿润,表皮细胞分泌水分、糖类、脂类、酚类、激素、酶等物质,可以粘住更多的花粉,并为花粉萌发提供必要的基质,这类柱头称为湿柱头,烟草、百合、苹果、豆科等植物的柱头属此类型;也有柱头是干燥的,在被子植物中较为常见,这类柱头在传粉时不产生分泌物,但柱头表面存在亲水性的蛋白质薄膜,能从薄膜下角质层的中断处吸收水分,所以在生理上这层薄膜与湿柱头的分泌相似,十字花科,石竹科植物和凤梨、蓖麻、月季等的柱头是干柱头,禾本科植物中的水稻、小麦、大麦、玉米等的柱头也属此类型。
(2)花柱花柱是柱头和子房间的连接部分,也是花粉管进入子房的通道。一般植物的花柱细长,如玉米的花柱特别细长;但也有极短不明显的,如水稻、小麦。多数植物的花柱中央为引导组织(transmitting tissue)所充塞,构成这组织的细胞长形、壁薄、细胞内含丰富的原生质和淀粉,常成疏松状排列,棉、烟草、番茄、荠菜以及大多数双子叶植物的花柱是这样的,也有的花柱中央是空心的管道,称花柱道(stylar canal),管道的周围是花柱的内表皮,或为 2—3层分泌细胞,如单子叶植物百合科的百合、贝母和双子叶植物的罂粟科、马兜铃科、豆科等。不论是哪一种情况,当花粉管沿着花柱生长并进入子房时,花柱能为花粉管的生长提供营养和某些趋化物质。花柱内有维管束分布,一端与花托相连,另一端止于柱头。
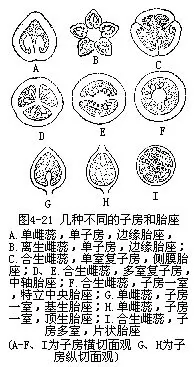
(3)子房 子房是雌蕊基部的膨大部分,有柄或无柄,着生在花托上。子房的中空部分称为室(Locule)。单雌蕊子房内仅有一室,如豌豆、牡丹等。复雌蕊的子房可由数个心皮合为一室或数室,如黄瓜的子房一室、烟草的二室,牵牛的三室,月见草的四室,凤仙花的五室等。有的植物子房原为一室,以后由于产生假隔膜而使子房分隔为多室的,如油菜、黄芪等。复雌蕊子房室数的差别,决定于各心皮的愈合状况以及心皮数目的不同。如果心皮彼此以边缘相连接,全部心皮都成为子房的壁,这样的子房是一室的。如果各心皮的边缘向内弯入,各心皮的边缘在子房中央部分彼此联合,心皮一部分成为子房的壁,而弯入部分的心皮成为子房内的隔膜,就成为多室。有的子房原为多室的,以后由于隔膜消失,成为一室,如石竹、报春花等(图4-21)。
子房的内、外壁上都有表皮、气孔和毛茸等结构。每个心皮有一条较大的中央维管束沿心皮的背缝分布,另有二条侧生维管束分布在心皮两侧的边缘,也即腹缝处(图4-20),这些维管束一方面分别向心皮输送水分和养料,同时也通过分枝向子房内壁的胚珠运送养料。
胚珠(ovule)是着生在子房内的卵形小体,是由心皮内侧若干部位的细胞经过快速分裂、生长后出现的突起所形成。子房内胚珠的数目视植物种类而异,常一至多个不等。着生的部位,一般在子房腹缝的一侧或在子房的中轴处,每一胚珠由珠心(nucellus)、珠被(integument)和珠柄(funiculus)所组成。
胚珠着生的心皮壁上,往往形成肉质突起,称为胎座(placenta)。由于心皮数目的不同,以及心皮连结的情况不一样,所以胎座有以下几种不同的类型:一室的单子房,胚珠沿心皮的腹缝线成纵行排列,称为边缘胎座(marginal placenta),如豌豆、蚕豆;一室的复子房,胚珠沿着相邻二心皮的腹缝线排列,成为若干纵行,称为侧膜胎座(parietal placenta),如罂粟、紫花地丁;复子房如分隔为多室,胚珠着生于各室的内隅,沿中轴周围排列,称为中轴胎座(axialplacenta),如水仙、百合、鸢尾等。多室复子房的隔膜消失后,胚珠着生在由中轴残留的中央短柱周围,称为特立中央胎座(free central placenta),如石竹、马齿苋等。特立中央胎座的短柱也有是心皮基部和花托上端愈合,向子房中央伸长而成的。此外,胚珠也有着生在子房基底的,如向日葵,称为基生胎座(basal placenta),或着生在子房顶部而悬垂室中的,称为顶生胎座或悬垂胎座(pandulous placenta),如桑。如果在多室子房中胚珠着生于隔膜的各面,则称片状胎座(lamellate placenta),如芡(图4-21)。
一朵具备以上各部分结构的花称为完全花,如果有一部分或二部分缺少不全的,称为不完全花。花萼和花冠全缺的称无被花,仅有花萼或花冠的为单被花。雌蕊和雄蕊如果在一朵花上同时兼备的称为两性花,单存一种花蕊而缺乏另一种的称为单性花,其中只有雌蕊的称雌花,只有雄蕊的称雄花。花被保存而花蕊全缺的称无性花或中性花。无被花、单被花、单性花和中性花都属不完全花。雌花和雄花生于同一植株上的,称为雌雄同株,分别生于二植株上的,称为雌雄异株。同一植株上,两性花和单性花都存在的,称为杂性同株,如槭、柿等。
(三)花各部分的演化
每一种植物,花的各部分形态是比较固定的,而花的形态变化又往往与植物的演化有关,因此,被子植物的分类依据,很大一部分是由花的形态来决定的。花各部分的演化趋势,主要从以下几方面表现出来:
1.数目的变化 组成花的各部分,在数目上是有不同的,总的演化趋势是从多而无定数到少而有定数,如玉兰、莲、毛茛等较原始植物的花,雄蕊、雌蕊、或花被的数目是多而无定数。而大多数被子植物的花,这三部分的数目有显著减少,但减少情况,各轮并不完全一致,一般减少到3数、4数和5数,或是3,4,5的倍数。一般说来,单子叶植物多为3数、或3的倍数;双子叶植物多为4数、5数,或是4,5的倍数。多数植物的雌蕊心皮数目常较花被为少,而雄蕊则比较增多。花部的相对固定数目(3,4,5)称为花基数。
花各部分数目上的关系,一般服从于花基数或它的倍数,例如石竹属植物的花基数是5,具5个萼片,5个花瓣,10个雄蕊,5个心皮,但也有和这个原则不符合的。另外,花部的数目在发展过程中有趋向于退化减少,甚至少于原基数的情形。如十字花科的花基数是4,按雄蕊2轮应为8个,可是有2个已经退化消失,只存6个,作二轮排列;再如紫丁香的花基数为4,而雄蕊中只存2个,其他2个也在演化过程中消失。部分退化的情形,除上述雄蕊外,也可见于其他花部,如花冠退化,仅留花萼。整个花被退化,仅存花蕊。或是二种花蕊中有一种退化的(单性花),又如雌蕊心皮数的部分退化(少于原基数),更是习见。
2.排列方式的变化 花的各部分在花托上的排列,是随植物种类不同而异的。花部的排列主要有两种方式:一种是螺旋排列;另一种是轮状或圆周排列,二者中,前者是原始类型。螺旋排列的花,从最外面或最下层的苞片起,继而花被,雄蕊群到中央的雌蕊群,在花托周围呈现由下而上,或由外而内的按顺序螺旋排列,双子叶植物中的毛茛科、木兰科等植物,仍保留这种排列方式。轮状排列就不是这样,花的各部分常由下而上,或由外而内地按顺序排列成一轮或数轮,每一轮的各个分体,常与相邻的内轮或外轮的各分体相间隔排列,也即萼片和花瓣相间隔,花瓣和雄蕊相间隔,心皮和雄蕊相间隔。有些花的结构看起来似乎不尽如此,例如鸢尾的心皮与雄蕊相对而生,报春花的雄蕊和花瓣相对而生,产生这类现象的原因,是由于某一轮花部的消失所致,如鸢尾与报春花原来各有2轮雄蕊,以后鸢尾的内轮雄蕊消失,保留下来的外轮雄蕊就和心皮对生。同样,报春花的外轮雄蕊消失,而内轮雄蕊就与花瓣相对而生。
此外,花部作螺旋排列的花托往往凸起或呈圆柱状,而轮状排列的花托多为平顶或凹顶。
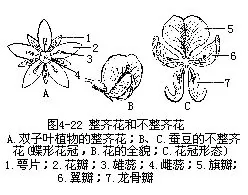
3.对称性的变化 花各部分在花托上的排列,常形成一定的对称面。如果通过花的中心可以作出几个对称面的,这类花称为辐射对称,也称整齐花。如果通过中心只可作一个对称面的,称两侧对称,这类花称不整齐花(图4-22)。桃、李和百合等花是整齐花,它们的花基数各为5和3,各轮花部都按相互间隔的规律和辐射对称的位置在花托上排列。蚕豆或其他豆类花的基数是5,5片萼片相互联合(仅上部分离),5片花瓣的大小、形状也极不相同,上方1片大形花瓣称旗瓣,侧面2片略小,形状相同,状如双翼,称翼瓣,另2片更小,合生在一起,位于花冠下方,形状如龙骨突起,故称龙骨瓣。花冠以内的雄蕊群共有雄蕊10枚,9枚的花丝合生在一起,另1枚单独离生。心皮数是一片。整个花冠外形似蝶,称为蝶形花冠,是两侧对称的不整齐花。也有完全不对称的花,如美人蕉,其花被虽属整齐型,但因雄蕊的部分退化和瓣化而使整个花朵显示不对称状,故一般常以此作为不对称花的例子。从进化的观点看,辐射对称是原始性的,而两侧对称,则是进化的。花冠的形状和对称性也往往与传粉方式有相关性,也是长期适应所产生的结果。
4.子房位置的变化 原始类型的花托是一个圆锥体或圆柱形,在进化的演变过程中,花托逐渐缩短,加大宽度,变为圆顶或扁平状,并且进一步在中央出现凹陷,成为凹顶形。花托形状的变化,改变了花部在花托上的排列地位,特别是子房的位置,出现下述几种不同的状态。
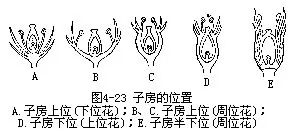
第一种是花托圆柱形,或是圆顶、平顶状,花萼、花冠和雄蕊群顺序地着生在雌蕊下方的花托四周,或是雌蕊外方的花托上,雌蕊的位置要比其他各部分高,象这样的子房位置称子房上位。子房上位的花也称下位花(hypogynous flower),如芸苔、毛茛、牡丹、蚕豆等(图4-23,A)。
第二种是花托中央凹陷,花托杯状或盂状,花萼、花冠和雄蕊群着生在杯状花托隆起的四周边缘上,而雌蕊的子房着生在花托的杯底,花托的壁与子房壁并不相连。这类子房的位置同样是上位。称这类花为周位花(perigynous flower),如蔷薇、月季、樱花等(图4-23,B、C)。
第三种是花托呈深陷的杯状,子房着生在花托的杯底里,子房的壁和花托完全愈合,只留花柱和柱头突出在花托外面,花萼、花冠和雄蕊群着生在子房上方的花托边缘上。这类花的子房位置最低,所以是子房下位,而称这样的花为上位花(epigynous flower),如梨、苹果、黄瓜等(图4-23,D)。
另有一种是花托呈深陷的杯状,子房着生在花托杯底里,只有子房壁的下半部与花托愈合,其余部分与花托分离;花萼、花冠和雄蕊群围生在子房上半部的周围。这类子房的位置为半下位(half inferior),也把这类花称为周位花,如忍冬、接骨木,虎耳草等(图4-23,E)。
花各部分的演化趋势是多方面的,就一朵花来说,演化的趋势也不是各部分同步一致的,这就使花的结构更为复杂而多样化。例如苹果和梨的花萼、花冠离生,雄蕊多数,这些反映了它们的原始性状,但凹陷的花托和下位子房,则又是进化的表示。此外,花各部分的互相转变,也经常在栽培的植物种类中找到,如栽培的芍药和玫瑰,雄蕊的数目减少,而花瓣增多,出现重瓣的结构。在睡莲的花朵里,也常常可以见到花瓣和雄蕊之间的过渡类型,这种瓣化雄蕊的下部花丝成为扁平,接近于花瓣的薄片状,而在上部却留有残存的花药(图4-24)。
