��һ�� ǻ�������ŵ���Ҫ����
��
����������ڶ����ݻ�����һ����֧��ǻ���������������������Ŀ�ʼ�����ද���ڽ���������ռ����Ҫλ�ã����������������ﶼ�Ǿ�������η�չ�����ġ�ǻ������Ϊ����Գơ������߲㡢����֯�ֻ���ԭʼ������ǻ��ԭʼ��ϵͳ�ĵ͵Ⱥ������metazoa����
������һ������Գ� ���������Ͷ����Dz��ԳƵġ���ǻ�����↑ʼ���������˹̶��ĶԳ���ʽ�����Ŷ���һ��Ϊ����Գƣ�radial symmetry�����������ǻ�����ͨ�������ڵ������ᣨ�ӿ��浽�����棩�������������������Ϊ2����ȵIJ��֡�����һ��ԭʼ�ĵͼ��ĶԳ���ʽ�����ֶԳ�ֻ���ϡ���֮�֣�û��ǰ������֮�֣�ֻ��Ӧ����ˮ��Ӫ���ŵĻ�Ư����������������ԳƵ����ٴ���Χ��������ȡʳ�����ܴ̼�����ǻ����������Щ�������ɷ���ԳƷ�չΪ������Գƣ�biradial symmetry������ͨ������������ᣬֻ������������������Ϊ��ȵ������֡����ǽ��ڷ���Գƺ�����ԳƵ�һ���м���ʽ��
�������������߲㡢ԭʼ����ǻ �������Ȼ���ж��߲㣬���ӷ������������������������ﲻͬ�����һ��ֻ��Ϊ����ϸ����ǻ��������Ǿ����������߲㣨�ڡ����߲㣩�Ķ���ڶ��߲�֮�������ڡ����߲�ϸ�����ڵ��н��㡣�������߲�ϸ����Χ�ɵ����ڵ�ǻ������̥�����е�ԭ��ǻ�����뺣�������ǻ��ͬ�����������Ĺ��ܣ�������ϸ���⼰ϸ������������ˣ�����˵�����ද�↑ʼ��������ǻ����������ǻ�ּ���ѭ�������ã����ܽ��������Ӫ���������͵���������֣������ֳ�Ϊ����ѭ��ǻ��gastrovascular cavity�����пڡ�û�и��ţ�������IJ������ɿ��ų������Ŀ�����ʳ����Dz�Ĺ��ܡ��ڼ�Ϊ��̥����ʱ��ԭ�ڣ���ߵȶ���Ƚϣ�����˵ǻ�������൱�ڴ���ԭ���߽Ρ�
������������֯�ֻ� ���ද����Ҫ����ϸ���ֻ���ǻ�����ﲻ����ϸ���ֻ������ҿ�ʼ�ֻ�������֯���������֯һ���Ϊ��Ƥ����ޡ����⡢�����࣬����ǻ��������Ƥ��֯ȴռ���ƣ������γ����ڡ�����棬���ֻ�Ϊ�о�ϸ��������ϸ���ȡ������ص�������Ƥϸ���ڰ����м�ԭ��ά������ϸ��������Ƥ�ͼ���Ĺ��ܣ����Գ�Ϊ��Ƥ����ϸ����epithelio-muscular cell�����Ƥ��ϸ����ͬʱǻ���������Ƥ����������һ���Ĵ������ܣ����ǽ�Щ����Ӧ�õ�����ѧ�����͵������������о�ǻ��������һ�����֡����Ĵ�����non-nervous conduc-tion��������neuroid����������������ǻ������õ�֤ʵ�ġ�
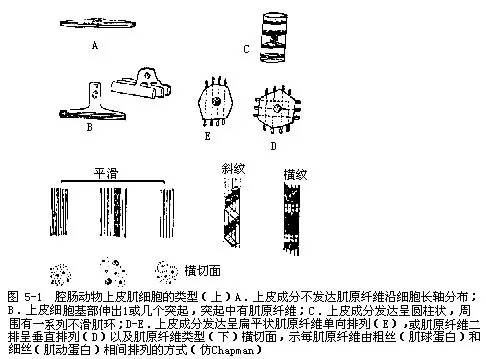
�������ģ�����Ľṹ��musculature�� ��Ƥ����ϸ����������Ƥ��Ҳ���ڼ���ķ�Χ���������Ƥ�뼡��û�зֿ�����һ��ԭʼ������һ������Ƥ����ϸ���Ļ��������һ����ϸ����ͻ�������м�ԭ��ά��myofibrils����ͼ5-1B����Ҳ�е���Ƥ�ɷֲ������Ϊ��ϸ����myocyte����ͼ5-1A�����е�����Ƥ�ɷַ��ϸ���ʱ�ƽ״����ԭ��ά�ʵ������У�ͼ5-1E����������2�ż�ԭ��ά�ʴ�ֱ���У�ͼ5-1D����Ҳ�е���Ƥ�ɷַ����Բ��״����Χ��һϵ�е�ƽ��������ͼ5-1C��������άҲ��Ϊ���Ƽ���б�Ƽ���ƽ������ͼ5-1����ÿ����ԭ��ά������һ��ϸ˿��ɣ���Щ˿�ִַ֡�ϸ2�֣���ߵȶ���֣����ף���ϸ���������ף�˿���ƣ�����������Ҳ�ߵȶ�������ơ����ڼ������֧���˽�IJ��࣬�������е�ʵ��֤����ǻ����������뼡��ĽӴ�����——����ͻ����neuromuscular synapses���ij��ṹ���������ӣ�neuromus-cular junction����Ҳ����ߵȶ�������ơ�
��
��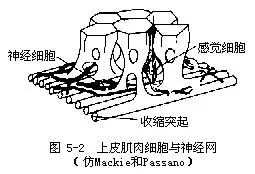
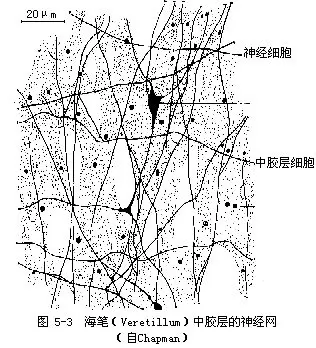
�������壩ԭʼ����ϵͳ——������nerve net�� �Ƕ�����������ԭʼ����ϵͳ��һ����Ϊ�����������ɶ����Ͷ༫����ϸ����ɡ���Щϸ��������̬�����Ƶ�ͻ��������γ�һ�����ɵ�������˳���������Щ����ֻ��һ�����������������߲�Ļ�����ͼ5-2������Щ������2���������ֱ�������ڡ����߲�Ļ���������Щ���������߲�������⣬���н�����Ҳ��������ͼ5-3������ϸ��֮������ӣ�����������֤����һ������ͻ�������ӣ�Ҳ�з�ͻ�������ӡ���Щ��ϸ�������ڡ����߲�ĸо�ϸ����Ƥ��ϸ��������ϵ���о�ϸ�����ܴ̼���ͨ����ϸ��������Ƥ��ϸ���ļ���ά�����������������ֽ���γ�������ϵ��neuro-muscular system�������������̼����⡢�ȡ���ѧ�ġ���е�ġ�ʳ��ȣ�������Ч�ķ�Ӧ���粶ʳ���ܵ��Լ�Э������Ļ�ȡ���ǻ������û�������࣬�Ĵ���һ��������ģ���˳�Ϊ��ɢ��ϵͳ��diffuse nervous system����ͬʱ���Ĵ����ٶ�Ҳ�����������˵������ٶ�Լ��1000�����ϣ��ⶼ˵��������ϵͳ��ԭʼ�ԡ�
��
������������ǻ��������ͻ��ij��ṹ���о������������ӵ�ͻ��������̬���м���������ֻ�����ӵ�һ��ͻ�������ݣ�����һ��û�С���û�м�����ͻ���ϣ�2��ͻ�����ݡ�������̬�ϵļ��������Ǵ���ϵͳ�м��������Ļ�����
��
�ڶ��� ǻ�������Ŵ�������——ˮHydra��
��
����ˮ������ڵ�ˮ�У���ˮ���ϻ�ˮ�ݷḻ����ˮ�г��ɲɵ���ˮ�ֲ��Ϲ㡢���ײɼ����������ұ��ڹ۲���ṹ����˳�����ʵ����ϡ�ͨ�������˽����ද��Ļ����ṹ��
����ˮ���ΪԲ��״���������������̼�ʱ�ɽ���������һ�š�һ�˸���ˮ�ݻ����������ϣ����Ŷ˳�Ϊ���̣�basal or pedal disk������һ���пڣ��ڳ���Բ�ε�ͻ��——������hypostome���ϣ�ƽ���ڹرճ����Σ�����ʳʱ���ſ����ڿ�֮��Χ����ϸ���Ĵ��֣�tentacle����һ��6��10�����ʷ������У���ҪΪ��ʳ���٣�ͼ5-4������ˮ��ʱ��������úܳ���������һ��������ʳ����ɴ����������͵����С�Ҳ�ɽ����ڴ��ֺ�����������������˶���˶���ͼ5-5����
��
��
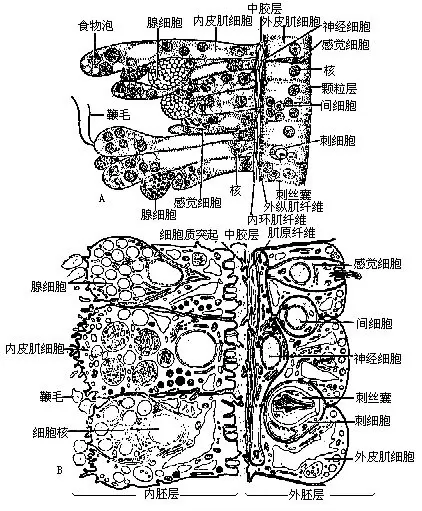
����ˮ�������ڲ�Ϊһ��ǻ���ɿ��������ͨ��Ҳ�봥����ͨ����Ϊ����ѭ��ǻ�������������ϸ�����ɣ���2��ϸ��֮��Ϊ�н��㡣�����һ��Ϊ���߲㣬���ϸ����Ҫ�б����о��Ĺ��ܡ������һ��Ϊ���߲㣬��Ҫ��Ӫ�����ܣ�ͼ5-6����

��
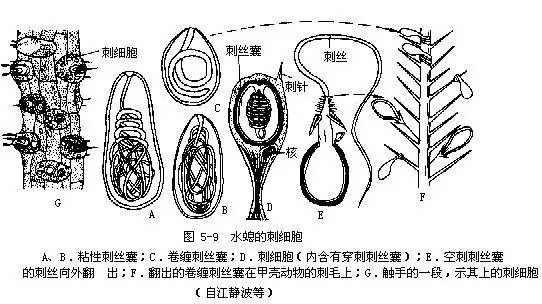
�������߲����Ƥ��ϸ��������Ƥ��ϸ������Ƥ��ϸ��������ϸ�����о�ϸ������ϸ������ϸ���ͼ�ϸ����ͼ5-7���������߲���Ƥ��ϸ����Ŀ��࣬Ƥ��ϸ�������ļ�ԭ��ά��������֮�������У���һ�����еļ���ά��������ʱ��ʹˮ�������ֱ�̡��о�ϸ����sensory cell����ɢ��Ƥ��ϸ��֮�䣬�ر��ڿ���Χ�����ֺͻ����Ͻ϶࣬�������С��ϸ����Ũ���˲��ио�ë������������ά���ӡ���ϸ����nerve cell��λ�����߲�ϸ���Ļ������ӽ����н���IJ��֣���ϸ����ͻ��˴����������γ���״��ͼ5-8���������̼���������ɢ�����Ե��������һ�����ܽ�ǿ�Ĵ̼�ʱ��ȫ��������������Ӧ���Աܿ��к��̼�����ϸ����cnidoblast����ǻ�����������еģ����鲼��������������ر�ࡣÿ����ϸ����һ��λ��ϸ��֮һ�࣬������״�Ĵ�˿�ң�nematocyst�����������ж�Һ��һ������˿״�ܡ�ˮ������ִ�˿�ң�ͼ5-9�������̴�˿�ң�������һ��ϸ���пյĴ�˿�����ܴ̼�ʱ����˿���ⷭ����������ָ�˴������ⷭһ�����ͿɰѶ������벶���������С�������ڣ�����������ɱ����������˿�ң���ע�䶾Һ����ֻ���Ʊ��������2��ճ�Դ�˿�ң��Բ�ʳ���˶������á���ϸ����interstitial cell������Ҫ�����߲�ϸ��֮�䣬��һ�Ѷѵ�Сϸ������С��Ƥ��ϸ���ĺ˲�࣬һ����Ϊ����һ��δ�ֻ�����̥�Ե�ϸ�������Էֻ��ɴ�ϸ������ֳϸ���ȡ���ϸ����gland cell������������У��Ի��̺Ϳ���Χ��࣬�ܷ���ճҺ����ʹˮ���������ϻ������ϻ��С�Ҳ�ɷ������壬��ճҺ����һ���ݣ�ʹˮ���ˮ��������ˮ�档
 ��
��
��
 ��
��
 ��
��
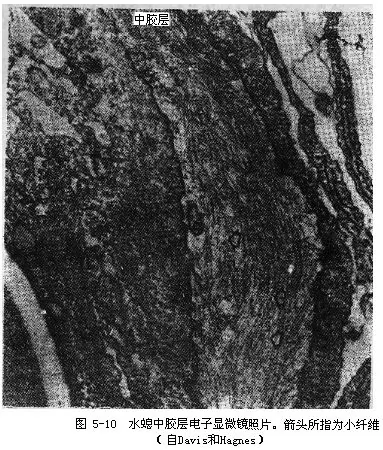
�����н��㱡������Ϊ�����߲�ϸ�����ڵĽ�״���ʣ�������ʹ��ֶ��������ġ��ڵ��������£��н������кܶ�С��ά��Ƥ��ϸ��ͻ��Ҳ�������У�ͼ5-10�����н��������е��ԵĹ�������������֧�����á�
�������߲������Ƥ��ϸ������ϸ���������о�ϸ�����ϸ����ͼ5-7���������߲�ϸ���Ļ���Ҳ�з�ɢ����ϸ������δ���ӳ�������Ƥ��ϸ�����Ӫ������ϸ����nutritivemus cular cell������һ�־�Ӫ�����ܼ��������ܵ�ϸ������ϸ��֮����ͨ����2����ë��1��5���������ڱ�ë�İڶ��ܼ���ˮ����ͬʱҲ�����α����ʳʳ�ϸ���ڳ����в���ʳ���ݣ�������ļ�ԭ��ά�������������֮���ijʻ������У�����ʱ����ʹ������ֱ�ϸ���ڿ���Χ��Ƥ��ϸ���ļ�ԭ��ά������Լ�������á���ϸ������Ƥ��ϸ��֮�䣬��ɢ�����߲�����֡���ϸ�������IJ�λ��ͬ���书��Ҳ��һ�������ڴ������ֵĿɷ���ճҺ���ڻ������ã�ʹʳ�����ױ��̽�ȥ��������ѭ��ǻ�ڵģ����ܷ�������ø����ʳ�
����ˮ��Ը���С�Ƕ������ࡢ��ˮ��ȣ���С�����׳��С���ڶ����Ϊʳ���Դ��ֲ�ʳ��������ʳ��ɱ�ˮ��ܶ�����ֽ�����������ڲ������ڲ������ܴ�˿�����˷ų������ģ�glutathione�����ڸ����ʵĴ̼��£�ˮ���ſ���ʳ���������ѭ��ǻ�������Ʒ�����ķ���ʢ��ˮ��ˮ�У���ʹû�в����ˮ�Ҳ���ֳ�ȡʳ�Ķ�����������ǻ�ڣ�����ϸ������ø����ҪΪ�ȵ���ø������ϸ����������extracellular digestion�������������γ�һЩʳ�����������Ƥ��ϸ���������ϸ����������ʳ�����ϸ�����������������ʳ��ɴ��������߲�ϸ������ɢ������ϸ�������������IJ����پ����ų����⡣
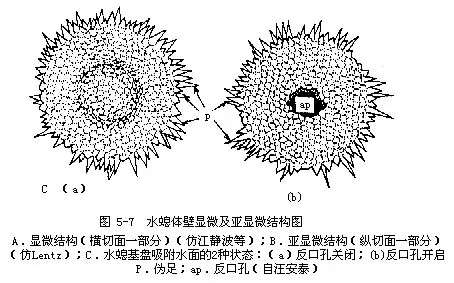
����������һ����Ϊǻ���������ˮ��ֻࣩͨ�����������ͨ����������ѧ�߷���ˮ�����Hydra������ˮ�����Pelmatohydra������ˮ�����Chlorahydra�������࣬�ڻ���������һ���ڿף�aboral pore�����۲�ˮ���̵ij��ṹʱ���֣���ԭ��ά�Է��ڿ�Ϊ���ijʶ������״���С��������з�ʽ�����ڿ��Ʒ��ڿĿ����رա����ڿ�ֹʱ�����̸��Ų�������Ŀ���������ͼ5-7C�������ڿ���ʱ���з��������ӿ����ų�������ˮ�ķ��ڿ��и��ŵIJ����������ܡ�
������������й��û����������٣��ɸ�ϸ���������ų�������̼�ͷ��
����ˮ����ֳ�����Ժ��������֡���������������ֳ——��ѿ��ֳ�����������ͻ���������γ�ѿ�塣ѿ�������ѭ��ǻ��ĸ����ͨ����ѿ�峤���������ںʹ��֣�������������ĸ�����룬��������Ӫ�������������ֳ�Ǿ��ѽ�ϡ����������Ϊ�������壬����Ϊ����ͬ�塣��ֳ���������߲�ļ�ϸ���ֻ��γɵ���ʱ�Խṹ������ΪԲ�Σ��ѳ�Ϊ��Բ�Σ�ͼ5-11�����ѳ���һ��ÿ�γ���һ���ѣ�Ҳ�е�����һ�γ��켸���ѡ����ڳ���ʱ���ѳ����ѣ�ʹ��¶�����������γɺܶྫ�ӣ�����ľ��ӣ����������ν�������֮�ܾ����ܾ��ѽ�����ȫ���ѣ��Էֲ㷨�γ�ʵ��ԭ���ߡ�Χ����̥����һ�ǣ���ĸ������������������ˮ�ף��ɹ��϶���ɺ�������������������תʱ����̥����䷢���������ѣ���̥�ݳ���������Сˮͼ5-12����


��
����ǻ�����������������ǿ�����ˮ��гɼ�С�Σ�ÿ�ζ��ܳ���һ��Сˮ����ֻ�е����Ĵ��ֲ��������������Ķ��ͨ��ˮ�Ĵ����Ϳ��п����ܳ���˫ͷˮͼ5-12�������������˽�ˮHydra oligactis��H.pseudoligactis�������߲�ϸ���ֿ������2��ϸ��������������Ϊ������ˮ��ˮ�������ͳ�ѿ���棬��ȥ��Ϊ��ϸ������Ҫ�IJ���ȱ�ٵ����ã��ɼ�ϸ���ֻ�Ϊ�������͵�ϸ��������������Ϊ����ϸ���������ͳ�ѿ�����в��Dz���ȱ�ٵģ�����X������������nitrogen mustard�������ƻ���ϸ�������ܽ����������ѿ��

��
������ ǻ�������ŵķָ�
��
����ǻ������Լ10000���֡���Ϊ3�٣�ˮ�١���ˮĸ�١�ɺ���١�
��
һ��ˮ�٣�Hydrozoa��
��
�������ٶ��������������ں�ˮ�У����������ڵ�ˮ������ʷ�д���ˮ��ͺ�ˮĸ�ͣ�������������������֦�棨Obelia����ͼ5-14����

����֦��������dz���������ں��塢��ʯ�����������ϣ�Ϊһ��֦״��ˮ���Ⱥ�壨ͼ5-13����Ⱥ������Ĺ������ֲ��ĸ����ʳ�����hydrorhiza����������������ܶ�ֱ���ľ�����Ϊ��hydrocaulus�����Ϸֳ�2�ָ���——ˮ��壨hydranth������ֳ�壨gonangium��������Ⱥ�����棬��Χ�������߲���ڵ�һ�����Ľ���Ĥ����Χ�ʣ�perisarc�����߱�����֧�ֵĹ��ܡ�ˮ�����Ҫ��Ӫ�����乹����ˮ������ͬ���пڼ����֣�������ʵ�ĵģ�������ˮ�ij���������һ���ı����ʣ���Ϊˮ��ʣ�hydrotheca������ֳ���ڼ����֣�ֻ��һ�пյ��ᣬ��Ϊ�Ӿ���blas-tostylus�����Ӿ�����Χ��������ƿ״�ʣ���Ϊ��ֳ�ʣ�gonotheca������ֳ������������ֳ����Ӫ����Ҫ��ˮ��幩������Ϊˮ������ֳ��˴����еĹ��⣨coenosarc�����ӣ�����Ⱥ�������ѭ��ǻ����ͨ���ġ�Ⱥ������һˮ��岶ʳ������ͨ������ѭ��ǻ�����������ֻ��������塣
������ֳ�������Ӿ��Գ�ѿ�ķ�����������ˮĸѿ��ˮĸѿ���죬�����Ӿ�������ֳ�ʶ��˵Ŀ��ڳ������ں�ˮ��Ӫ���������ṹ�ϼ���С��Լ1mm��2mm��������һԲɡ��ɡ��Ե���кܶ�ϸ�Ĵ��֣�����ʱ16��������ɡ��������һ�̵Ĵ�����������ͨ��θ������θ���4�����ܣ���ɡ��Ե�Ļ�����ͨ���ڡ�θ�����ܡ����ܹ���ˮĸ������ѭ��ϵ��ˮ�ˮĸ����֮һ����ɡ�����Ե��һȦ��Ĥ��ΪԵĤ��velum������֦���ˮĸԵĤ�˻�����ɡ��Ե��8��ƽ���ң�˾ƽ�⡣��4�����������ĸ������߲��γɵľ������ѳ����������壩�����ѳ�����ں�ˮ���ܾ����ܾ��ѷ����������Ƶķ�ʽ�γ�ʵ�ĵ�ԭ���ߣ��������������ë�����ζ�����Ϊ�����׳棨planula���������׳��ζ�һ��ʱ�ں����������Գ�ѿ�ķ�ʽ������ˮ��͵�Ⱥ�塣
��
����֦�������ʷ����2���Ρ�ˮ���Ⱥ�������Գ�ѿ�ķ������������ˮĸ�ͣ�ˮĸ������������ֳ��������ˮ���Ⱥ�壬�������λ��ཻ�棬����������������ʷ��
������֦��Ľṹ������ʷ�ɿ������ٶ������Ҫ������
����1.һ����С�ε�ˮ��ͻ�ˮĸ�Ͷ��
����2.ˮ��ͽṹ�ϼ�ֻ�м�����ѭ��ǻ��
����3.ˮĸ����ԵĤ�����ֻ�����ƽ���ҡ�
����4.����ʷ����ˮ�����ˮĸ�ͣ�������������������֦�棩����������ˮ��ͷ����ˮĸ�ͣ���ˮ��ˮĸ�Ͳ������Ͳ�Tubularia����Ҳ��ˮĸ�ͷ��ˮ��Ͳ�������ڣ��繳��ˮĸ��Gonionemus�����һ�ˮĸ��Craspedacusta�������е�Ⱥ�巢չΪ��̬������ɮñˮĸ��Physalia����ͼ5-14��15����

��
��
������ˮĸ�٣�Scyphozoa��
��
�������ٶ���ȫ�������ں�ˮ�У����Ϊ���͵�ˮĸ�ࣨ����һ��ϼˮĸ Cyanea arcticaɡ��ֱ�������2m�࣬���ֳ�30��m����ˮĸ�ͷ��ˮ��ͷdz��˻����������׳����ʽ���֣�����ˮĸ�͵Ĺ����ˮ�ˮĸ���ӡ��纣��ˮĸ��
��������ˮĸ��Aurelia aurita Lamarck����ͼ5-17��ӪƯ�������Ϊ��ɫ������״����ɡ�ı�Ե���д��֣�����8��ȱ�̣�ÿ��ȱ������һ���о�����Ҳ�ƴ����ң������и��ʵ�ƽ��ʯ��stato-lith�������������۵㣨ocellus������������Ե�꣨lappet����Ե�����ио�ϸ������ë��������2�����ѣ�ͼ5-18������ˮĸ�岻ƽ��ʱ�������ҶԸо���ë��ѹ����ͬ����������ƽ��ĸо�������ɡ��������һ���Ľ��εĿڣ��ɿڵ��Ľ������ 4������oral lobe��������ѭ��ϵͳ�Ƚϸ��ӣ��ɿڽ�ȥΪθǻ��λ�������룬���ķ�������ĸ�θ�ң���θ���Ϻ�θ��֮�������֦�ĺͲ���֦�ķ��ܣ�radial canal������Щ���ܾ���ɡ��Ե�Ļ��ܣ�ring canal��������ˮ���ɿڽ�ȥ��θǻ������һ���ķ��������ܣ�Ȼ������һ���ķ�������θ�ң�������������θ�ҵ����棬���ĸ������߲�����������ε���ֳ�٣�λ��θ�ҵײ��ı�Ե������ֳ���ڲ࣬���кܶ�˿״�Ľṹ��Ϊθ˿��Ҳ�������߲��γɵģ������кܶ��ϸ����ʳ�����θ�Һ�����˿ɱ������������ϸ���ں�ϸ�����������ɷ��ֲܷ���ȫ��������θ˿Ҳ���ű�����ֳ�ٵ����á�

��
��������ֳ�ٲ������ӻ��ѣ���ˮĸΪ�������壩�����ӳ������ˮ�����������ܾ���Ҳ�е��ں�ˮ���ܾ����ܾ��Ѿ���ȫ���������γ����ߣ��������ݷ�ʽ�γ�ԭ���ߣ���ʱ��̥���泤����ë����Ϊ�����׳棬�ں�ˮ���ζ�һ��ʱ�ں��ں�������������ϣ�������С���״���壨hydru-la�����пںʹ��֣���Ӫ�������Ȼ����к��ѣ��ɶ����·ֲ�Ϊ�������壨scyphistoma�����ٽ�������������γ�һ������״���壬�ƺ����壨strobila��������������һ��������������������Ϊ��״���壨ephyra��������������ˮĸ���塣�ɴ˿ɼ�����ˮĸ������ʷ��ͼ5��19�������������棬��ˮĸ�ͷ����ˮ������˻���Ϊ�׳档Ҳ�еIJ�ˮĸ��������������ֻ��ˮĸ�͡�
������ˮĸ��ˮ�ˮĸ����Ҫ��ͬ�����ڣ�
����1.��ˮĸһ��Ϊ����ˮĸ����ˮ�ˮĸΪС�εġ�
����2.��ˮĸ��ԵĤ����ˮ�ˮĸ��ԵĤ����ˮĸ�ĸо�����Ϊ�����ң�ˮ�ˮĸΪƽ���ҡ�
����3.��ˮĸ�Ľṹ�ϸ��ӣ���θ������θ˿����ˮ�ˮĸ���ޡ�

��
����4.��ˮĸ����ֳ����Դ�����߲㣬ˮ�ˮĸ����ֳ����Դ�����߲㡣
������ˮĸ����ǻ���������Ǿ��ü�ֵ�ϸߵ�һ�ද����纣�ؼ������ࡣ���أ�Rhopilemaesculentum����ͼ5-20���Ľṹ�뺣��ˮĸ��������һ�µģ���ͬ���ǣ����ص�ɡΪ�����Σ��н���ܺ��д�����ˮ�ֺͽ����ɡ�ı�Ե���֣���8���о�����ɡԵƽ��Ϊ8������ÿ������ɡԵ��14��20��С��״��Ե�꣨lappets������ĿΪ�ֵķ�������֮һ�����������ϣ����ο���ʧ���ڿڱ��Ļ�����8����״��壬�ڿڱ��²�����8������ÿ�������ֳַ������������Ե���γɺܶ�С�ף���Ϊ���ڡ��ڿ����ϳ��кܶഥ�֣����˿״����������״������������״�������������ͨ������ĩ����һ�����İ�״�������������Ҳ�кܶ����ڼ����֡����ؾ��ǿ�������ʳһЩС�Ķ�ֲ��Ϊʳ���������Χ�Ĵ����Ȱ�С������Ի�ɱ����Ȼ������ڣ��������кܶ��֦��С�ܵ�θǻ�����ֹ����ȡʳ������е���ֲ����������ϣ���˽�������һ��IJ�ˮĸ��ΪһĿ����Ϊ����ˮĸĿ��Rhizostomae����
�������ں��ص�����ʷ��ͼ5-21�����������ҹ���������ϸ�ı�������Է�ֳ����ѧ�ͷ�������ѧ���о����Ժ�����Դ��Ԥ��Ԥ������ֳ������Ҫ���塣
�������ص�Ӫ����ֵ�Ϸḻ�����е����ʡ�ά����B1��ά����B2�ȡ����ӹ����������Ƥ���Ǻ��ص�ɡ������ͷ����צΪ���صĿڱ����֡��ҹ�ʳ�ú��ص���ʷ�ƾã����ҹ��غ����صIJ����dz��ḻ���㽭�������غ�һ����ࡣ

��
�����������⣬������IJ�ˮĸ����ҵ�����к�������Σ�����㡢���࣬�����ƻ����ߡ��� 1952���ڸ��������غ�������һ�ֲ�ˮĸΪ�ֵ��������������߱��ƻ����²���������ǻ������Ĵ�˿�Ҷ��˵�Σ���ܴ���һЩ���ˮĸ�����������������ش��ˡ��Դ�˿����Ķ������ʵijɷ����ʣ��������ѽ�����һЩ�о���Ŀǰ�����о��ܶຣ������Ĵ�˿�Ҷ��أ���Ϊ�µ�ҩ����Դ����������ҽѧ�������1972����ѷ�����4��ǻ���������ȡ�����п�������ҩ�
��
 ��
��

��������ѧҲ���о�ˮĸ������Ԥ��籩�ı�����������ǰԤ�⺣�Ϸ籩Ҫ���״�վ��ˮ��վ�������������ǽ����ۺϹ۲⣬ʮ�ֲ��㣬�������ں�ˮ�е�ˮĸ�ڽ��������з�չ��һ��Ԥ��籩�ı���װ�ã�ʹ���ڷ籩����ǰ����Сʱ�������ģ��ˮĸ�о����ķ籩Ԥ����������ǰ15��Сʱ����Ԥ������ָ���籩���ķ���װ�ü��������㡣���纣�ص��˶���������ʽ��������ƽ��ģ�������ʽ�ɻ����������ϵ�����������ƽ��ġ��еĿ�ѧ��������Ѻ��ص��ƽ���ʽ��������ʽ�ɻ�����ƣ��������ܽ�ʡ������������õ������������Ķ�����

��
��
����ɺ���٣�Anthozoa��
��
������ٶ�����ǰ���ٲ�ͬ��ֻ��ˮ��ͣ�û��ˮĸ�ͣ���ˮ���Ĺ����ˮ�ٵ���帴�ӡ�ȫΪ��������������ů����dz���ĺ��ס�����“����”����ҪΪɺ���档һ����������ɺ��Ϊ��������غ�������Ϊ������
����������ͼ5-22�������������Բ��״��һ�˸��ں�����ʯ�����������ϣ��ö˳�Ϊ���̡���һ���пڣ����ѷ��Σ�����Χ���ֳ�Ϊ���̣�����Χ�м�Ȧ���֣��������д�ϸ���������Բ�ʳ��Ϻ�����С�����ʳ��ڣ�����ڵ���stomodaeum�����ڵ����ǿڲ������߲�ϸ�������γɵģ���Ϊ���������ڿڵ������˸���һ��ë���� siphonoglyphe����ƿڵ�������Щ����ֻ��һ����ë��������ë���ڱڵ�ϸ������ë��������������һ��ʱ��ˮ���Կ�����ë����������ѭ��ǻ��
��������ѭ��ǻ�Ĺ���ϸ��ӣ������п���խ��ͬ�ĸ�Ĥ��mesentery�������ɺܶ�С�ҡ���Ĥ������������߲�ϸ����������ͻ���γɵģ���2�����߲�֮��Ϊ�н��㡣��Ĥ�����ã���ҪΪ֧�֣�������������������ݸ�Ĥ�Ŀ��ȿɷ�һ������������ֻ��һ����Ĥ��ڵ��������ڸ�Ĥ����ı�Ե�и�Ĥ˿��mesenteric filament������Ĥ˿�ظ�Ĥ�ı�Ե���У�һֱ�ﵽ����ѭ��ǻ�ĵײ����еĴ�ײ�ʱ�γ��������״���Ϊ��˿�����к��зḻ�Ĵ�ϸ��������������ʱ�����ɿڻ�ڿ�������з����������Ļ��ܣ�ͼ5-23������Ĥ˿��Ҫ�ɴ�ϸ������ϸ�����ɣ���ɱ���������ڵIJ����������ϸ����������Һ����ϸ����������ϸ��������������Ϸ���ڽϴ�ĸ�Ĥ�϶���һ�ݼ������Ϊ���죨muscle band������Ĥ�ͼ���������Ƿ��������֮һ��

��
��������Ϊ�������壬��ֳ�ٳ��ڸ�Ĥ�Ͻӽ���Ĥ˿�IJ��֣������߲��γɡ����ӳ�����ɿ�������������һ���������ѽ���γ��ܾ��ѣ�Ҳ�е��ں�ˮ���ܾ�����ĸ���ڷ����γɸ����׳棬��ĸ�壬�ζ�һʱ�ں��������������¸��塣��Ҳ�еĺ������������׳棬ֱ�ӷ�����Ϊ�������ĸ�塣��ˮĸ�͡�������ֳΪ�ݷ��ѻ��ѿ��
�����Ӻ������˽�ɺ����Ļ����ṹ������ˮ�ٵ������IJ�ͬ�����ڣ�
����1.ɺ����ֻ��ˮ��ͣ��乹��ϸ��ӣ��пڵ����ڵ�������Ĥ��Ĥ˿��ˮ�ٵ�����幹��ϼ�ֻ�д������������ṹ��
����2.ɺ������������ֳ���������߲㣬ˮ����������ֳ���������߲㡣
���������ǵ���ģ��������ܶ�ɺ����ΪȺ�壬���߹�����
����ɺ���������γɴ����ɺ��������߲�ϸ���ܷ��ڹ������ڰ˷�ɺ���Ǹ٣�Octocorallia�������ֺ�Ĥ��8�����������߲��ϸ�������н����з��ڽ��ʻ�ʯ���ʵĹ�����Ƭ����Щ����������н����л�ͻ��������棬�纣���ڣ�Alcyonium���ͺ�����Pennatula�����е�����С��Ƭ���ӳɹ�״�Ĺ���������ɺ����Tubipora�������еĹ�����Ƭ���ϳ�����ǣ����ɺ����Cor-allium����ͼ5��23��ͼ5��24��������������ɺ���Ǹ٣�Hexacorallia�������ֺ�Ĥһ��Ϊ6�ı�������ʯɺ��Ŀ��Medreporaria���е�����Ⱥ�壬ÿ�������뺣�����ƣ�����̲�������ڵ����߲�ϸ���ܷ���ʯ�������ʣ������ڳ���ĵ��桢���漰��Ĥ��ȴ�������ÿ�����嶼����һ��ʯ�����ϣ���Ϊɺ������corallite����ͼ5-25������ʯ֥��Fungia����Ⱥ��ɺ�����乲�ⲿ�ֵ����߲�Ҳ����ʯ���ʣ�����Ⱥ�����״��ͬ�����������״Ҳ��һ�����е�Ϊ��֦״����¹��ɺ����Madrepora��Acropora�����е�ΪԲ��״������ɺ����Meandrina����ͼ 5-26����

����
����ʯɺ���Ĺ����ǹ���ɺ������ɺ��������Ҫ�ɷ֡��ɴ�����ɺ�������ѻ��ɵĵ��죬���ҹ�����ɳȺ����ӡ���������������̫ƽ���쳼�Ⱥ���ȡ��콸��ʯɺ�������߲�ϸ���г��д����ĵ�ϸ������壨Zooxanthella�����乲����������ɺ����ӳ���岹�����������࣬���ٹ������������������ɺ����Ĵ�л������CO2�����ȣ���������й�����ã���ʯɺ��������ϰ��Ҫ����ů��һ��Ҫ��ˮ��22�桫30�棩��dzˮ��ˮ��Լ��45m���ڣ��Ļ�������ˮ������һ���ij�������������ߵ�ɺ�����ܺ�ˮ����IJ����������ã��������Ź����Ķѻ��������ź������������ƣ�����չ���γɴ�ĵ��졣���غ��İ��������纣���ϵ���Ȼ���̣���ʹ������̡����ں��İ������ɷ������С�
��
��

����ʯɺ�����������Ƿ��ӣ��纣���غ�һ����ɺ������ķ��Ӽ�����á��������ۡ�������ʯɺ����ʯ����ˮ�ࡢ��·�ȡ��ҹ�̨�������ֵ�������ʯɺ���̳ɵģ�·����ƽ̹����������ֳʯ���ˣ����������á�����װ��Ʒ�ȣ���֮����ɺ������;�Ǻܹ�ġ�
����ɺ�������Եؿ��γ�Ҳ��һ�����á��ڵ����ϳ�����ʯ����ɺ�������γɵ�ʯ���ң�һ���Ϊɺ��ʯ���ҡ�������ʯ���ҵĵط���˵����������������ǰ��������ů��dz�������ҹ��Ĵ������������ǿ������Ԫ���������ʯ���ң���֤��������Ӧ��־���͡���ɺ�������ִ�ɺ�������γɴ��Ͳ㣬����Ѱʯ��Ҳ����Ҫ���塣
��
���Ľ� ǻ�������ϵͳ��չ

��
��
����ǻ��������������ϸ������Ŀ�ʼ��������巢������һ�㺣����ǻ����������������׳�ĽΣ��ɴ˿����룺��ԭʼ��ǻ���������ܹ�������Ӿ�ġ�����ë�Ķ������״�����׳棬��÷����·��������Ⱥ���ë�棬ϸ��������γ�Ϊԭʼ���߲�Ķ����չ��ǻ�����

��
�������ִ��ǻ�������У�ˮ����������͵ȵ�һ�࣬��Ϊ��ˮ�����ˮĸ�͵Ĺ��춼�Ƚϼ���ֳ���������߲㡣��ˮĸ��ˮ����˻���ˮĸ�ͷ���ṹ�ϸ��ӡ�ɺ������ˮĸ�ͣ�ֻ�нṹ���ӵ�ˮ��͡���2�ٵ���ֳ���ֶ��������߲㣬��˿�����Ϊ����ˮĸ�ٺ�ɺ���ٿ�����Դ��ˮ�٣����Ų�ͬ��;����չ�����ġ�
��������������ǻ�������ŵ����ݿɼ�Ҫ�������£�
����ǻ������һ��Ϊ����Գƣ�Ҳ���������ԳƵģ��������߲㣬��ԭʼ������ǻ������ѭ��ǻ�����п����ţ���ϸ���⼰ϸ��������������֯�ֻ�����ԭʼ�ļ���ṹ��Ƥ��ϸ������ԭʼ����ϵͳ�����������д�ϸ�����й���ʱ��Ϊ���ʻ���ʡ�����һ��Ϊˮ��ͺ�ˮĸ�ͣ�ˮ������ڹ���������ԲͲ�Σ�ˮĸ������Ư�������һ������Σ����ߵĽṹ������ͬ��ͼ5-27�������ˮ��͵��ã��������ˮĸ����ɡ�൱��������ѹƽ�������Ρ�����֮��ͬ�㣺ˮĸ����ӦƯ������н���Ӻ�����Լ�������ı��ء�Ҳ��ЩΪ��̬��Ⱥ�塣������ʷ�У���Щ�߷����ˮ�����ˮĸ�ͣ�����������������Щˮĸ�ͷ��ˮ��Ͳ�������ڣ���Щˮ��ͷ��ˮĸ�Ͳ�������ڡ�����������巢���о������׳�ʱ�ڡ���ȥһ����Ϊ��Щ�����о��ü�ֵ�����ſ�ѧ�ķ�չ������ǻ�����ﻹ�ж�����;��ֵ��̽����
��
������ˮĸ�����ţ�Ctenophora��
��
������ˮĸ��ȥ������ǻ�������ţ���Ϊ�̰����ţ�����Ϊ��ˮĸ�١�����һ���������Ϊһ�š�Ҳ���˰�����ǻ�������Ų��У�����Ϊ���䶯�
������һ�ද������ࣨ����100�֣����������Ƚ��٣�ȫ�������ں�ˮ�У��ܷ��⣬Ӫ�������Ҳ�е������С����������Ρ����Ρ���Բ���Լ���ƽ��״�ȣ�ͼ5-28������̬�ṹ��ǻ��������У����ͻ��������ڷ���Գƣ����������Գƺ����ԣ�����Ҳ���ڡ����߲㼰�н��㣻����ѭ��ǻ�벧ˮĸ�����ƣ����з�֦�ķ��ܣ�����֮�⣬����û������ǻ��

��
�������Ŷ���Ҳ��������������û�е��ص㣺
����1.��ˮĸ�������8�����е��ΰ壨combplate����ͼ5��29����ÿһ�ΰ�����һ�л�����������ë����ɡ��ΰ������м���άʹ�ΰ��˶����ΰ�Ϊ�˶�����

��
����2.�д��ֵ���ˮĸ�����������һ�����һ�ƴ����ʣ�tentacle sheath�������ڸ���һ�����֣�������û�д�ϸ����Euchlora rubra���⣬�д�ϸ������ճϸ���������д�����ճϸ����colloblast������ϸ���������ճ�����ʣ������Բ�ʳ��ϸ���ڲ�������״˿�������ﱻճ�ź���������������ϸ����ͼ5-30����

����3.�ڷ�������һ���еĸо����٣�sense organ�����ṹ�ϸ��ӣ�ͼ5��31������ƽ�����ڣ���4��ƽ����ë��֧��һ�����ʵ�ƽ��ʯ��statolith������ƽ����ë����������ë����ciliated furrow����8�����е��ΰ���������ˮĸ�ĸо�������˾ƽ������١�
��
��
����4.��ϵͳҲ�ϼ��У���Ȼ�����߲������������������8���ΰ弯�У��γ�8������������
����5.��̥�����У�����Ϊ�ѿ�ʼ���ֲ���������߲�ϸ����������չ�ɼ���ά��
���������Ͽ�������ˮĸ���ڽ�����Ϊ�����һȺ����ǻ������ӽ�������ǻ��������Ϊ�ߵȡ��е�ѧ����Ϊ������ˮĸ���ܽ���Ϊ���ζ����һ����Ϊ��ˮĸ���ڽ�������һä��֧������ߵȶ���û��ֱ�ӹ�ϵ��
������ˮĸ���Ը�������Ϊʳ��ͬʱ��������������Ķ��ϣ����������ʳ��������һ�����á���ˮĸ��ܳ�ĵ���׳桢���Ѻ����磬��ĵ����ֳ��ijЩ��ķ�ֳ��һ��Ӱ�졣
�������ද����Ȼ�������࣬���ǽ�����ä�ˣ���ȴ�����˿�ѧ���Ƕ�������Ȥ��ϣ�����������һЩһ������ѧ���⣬����ȡ��һ���ijɹ������������������ˮĸ��4�ִ���ϵͳ���ΰ�ϵͳ����Ƥ���������ΰ�����ϵͳ����ϸ����Ĵ���ϵͳ��Э�������ϵͳ����ϣ���Դ��������ԭʼ�Ĵ���ϵͳ��β�������4�ִ���ϵͳ�����Ժ���������������������β�������ߵȶ�������һ��������ϵͳ���ɼ���������ػ���С��Ⱥ���൱��Э��ϵͳȴ��ʾ������Ϊ���ƺͽ�����һ��ԭ����
��
��ϰ��
��
����1.ǻ�������ŵ���Ҫ������ʲô������������ڶ��������ռ��Ҫλ�ã�
����2.����ˮ�Ļ����ṹ�������߲�ϸ���ķֻ��ȣ�ͨ�����˽�ǻ���������ڽṹ����֯�ֻ��Ȼ���������
����3.ǻ��������ļ����٣����ٵ���Ҫ������ʲô���кμ�ֵ��
����4.�����˽�ǻ�������ϵͳ��չ��