��һ�� ��ٵ���Ҫ����
��
�������������������ë�����������º������ĸߵȼ����������ѧ�۵�������������ͻ�����������³´�л��ʢ�������ڿ����з��У���Ҳ������������������ĸ�������ʹ����������9ǧ���֣��ϳ�Ϊ���������࣬�鲼ȫ��ļ����
����������Դ�������࣬������ṹ���ܷ����кܶ���������������������������������ǹ��������ࣨSauropsida������������ͬ������ĸ����������������¼�����Ľ�����������
����1.���и߶��㶨�����£�ԼΪ37.0�桫44.6�棩�������˶Ի����������ԡ�
����2.����Ѹ�ٷ�����������ܽ�����Ǩ������Ӧ���Ļ���������
����3.���з������ϵͳ�й٣��Լ��������ϵ�ĸ��ָ�����Ϊ���ܸ��õ�Э�����������ͳһ��
����4.���н����Ƶķ�ֳ��ʽ����Ϊ���쳲�����Ѻ�����������֤�˺���нϸߵijɻ��ʡ�
����ѧϰ���������ṹ���ܣ�Ӧ������������Ϊ��������ע���ܽ�����������������Ƶ������Լ�����Ľ����������Ļ����ϣ��ص��������������Ӧ��������ʽ��������ṹ�������Լ����ʽ������������ػ���
��
һ�����¼����ڶ����ݻ�ʷ�ϵ�����
��
���������벸����Ǻ��¶�����Ƕ����ݻ���ʷ�ϵ�һ����Ϊ��Ҫ�Ľ������¼������¶�����нϸ߶��ȶ����³´�лˮƽ�͵��ڲ��ȡ�ɢ�ȵ��������Ӷ�ʹ���±�������Ժ㶨�ġ��Ը��ڻ����¶ȵ�ˮƽ�������������Լ��͵ȼ�������ࡢ�����ࡢ�����ࣩ���ű��ʵ����𣬺��߳�Ϊ���¶�����¶�����ȴ�л�����ǣ��³´�лˮƽ�ϵ͡����²��㶨��ȱ�����µ��ڵ�������
�����߶��㶨�����£��ٽ������ڸ���ø�Ļ�������̣�ʹ����ǧ�Ƶĸ���ø����Ӧ������Ļ�ѧЭ�����Ӷ����������³´�лˮƽ�����ݲⶨ�����¶���Ļ�������������Ϊ���¶����6�������˰Ѻ��¶������Ϊһ����ķ���Ͱ����˵�����Դٽ����ܴ�л��������塣�ڸ����£�����ϸ�����ر����ͼ���ϸ�����Դ̼��ķ�ӦѸ�ٶ��־ã������ճ�����½�������������������������������˺��¶�������˶��������������ڲ�ʳ���ܵС����»������˶���绷���������ԣ�����������ͷֲ��ķ�Χ���ر��ǻ����ҹ����������������¶���������һ����ҹ�䴦�ڲ��״̬���������͵����ں���������������Ϊ������������������֮������սʤ��½����ռͳ�ε�λ�����������Ҫԭ��
�������¶�������¾��Ը��ڻ����¶ȣ������������价���¶��£��л���ɢ�����ס��ڵ��ڻ����¶������������“����”�������������¶���������ֲ��ܹ��ߣ���������������������⣬�ܶ൰�����ڽӽ�50��ʱ�����ԣ�denaturation����
���������Dz��Ⱥ�ɢ�ȹ��̵Ķ�̬ƽ�⡣������ɢ���൱���������¼��ɱ�������ȶ���ʧȥƽ��ͻ��������²������������������������벸����֮������Ѹ�ٵص������Ⱥ�ɢ�ȣ�������и߶ȷ����������ϵͳ������صġ����µ������ࣨ�����²���ͨ�����ڷ����ٵĻ�����Э�����ɴ˿ɼ��������Ǽ���������ṹ����ȫ������IJ�����ȵ����ﻯѧ���ƵĻ��������ǣ�������ļ�״���������ڼ��⡢�κ����࣬��������ϸ��Ĥ���ϵġ�������Na+��K+��ATP�����������ᣩø��ʹATP�ֽ���ų�������
�������µij��֣��Ƕ����л����������ķ�չ�������뻷����������ͳһ�Ľ�������ݽ������Ĵ���ʵ��֤ʵ����ʹ�DZ��¶�����еĸ�������Ҳ��ͨ����ͬ�IJ���;����ʵ����ʱ�ġ����ڻ����¶ȵ����¡����磬��ң�⼼��֪̽��ijЩ������Ӿ�ĺ������ࣨһЩ��ǹ�㼰���㣩��ͨ������IJ��ȼ���Ⱥ���������ȣ��Լ����ӵ�ѪҺѭ��ͨ·��ʹѪҺ�������еĸߴ�л������������ѪҺ������Ѫ�ܶ�ɢʧ��ˮ�У����Ӷ���ø���ˮ�µ����¡���һ�������۳��������ң���������ˮ����10��仯��Χ��14�桫5�棩������£�θ���¶��Կ��ȶ���18�����ҡ�һ�ָ�ɽ���棨Liolaemus multiformis���ڽӽ������ϡ��������£��������Ϊ31�棬���ǽ�Ƥ������̫���ķ����ȶ�������µġ�һ��ӡ�����ߣ����߿ɽ����弡��IJ������������ȣ��Ȼ����¶ȸ�7�棩�Ӷ��������Ƶ��ѷ�������Щ��ʵ��һ��֤ʵ“һ�в��춼���м���ںϣ�һ�ж����������м价�ڶ��������”��
��
������ٵ�����ṹ
��
������һ��������������ʷĴ��Σ����ⱻ����ë��feather�������������͵��������Ӷ������˷����е�������ͷ�˾߽��ʵ��bill��������ʳ���١����״��ʳ�������й�ϵ����������β�˻������ɽ��ܼ�ʵ����֫ǿ����Щ������������ʽ������صģ����ɼ�ʵ��β���˻������ڷ��е��ȶ�������������ֲ�ǰ֫��ɳ���IJ��㣻�۴�������˲Ĥ�ɱ�������˲Ĥ��һ�ֽ�������Ĥ�����ڷ���ʱ�ڸ������Ա�����������ͻҳ���������˺�������˲Ĥ��Ե����һ����״��Ƥ��feather epithelium�����ڵ����Ե����ࣨ�������������Ϊ����ܽ���ˢϴ�ҳ���ˮ�ݼ���������٣�ͼ19—1�����������ݣ���Χ���������������ռ�������ҹ�������ࣨ��èͷӥ���Ķ���Ϊ���
����ǰ֫��Ϊ����wing������֫��4ֺ���������������������������ﲻͬ��������־��Ĵֺͨ���������������֦��������ֺ����̬�����ʽ�����й�ϵ��
����β����������״�����𣬳�Ϊβ���ڷ��������Ŷ�����á�β�����״������ص��йء�
����������Ƥ������Ƥ�����ص��DZ��ɶ���ȱ�����塣�����ɵ�Ƥ�������ڼ�������˶��������Ƥ��ȱ�����壬������������Ϊ���ơ�����Ψһ��Ƥ���ٳ�β֬�٣�oil gland�� uropygialgland�������ܷ��������Ա�����ë���±��Σ����ɷ�ˮ�����ˮ�ݣ�Ѽ����ȣ���β֬���ر����Щ���ࣨ������š����ҡ�𱼰���ĵȣ��ߡ����ķ�������һ����֬����ܻ�����ά����D���������β֬�ٵĻ�ѧ�ɷ��ڷ���ѧ�ϵ��������⣬������ѧ��������ӡ�Ҳ���˱������ڼ���������ȵ�Ƥ������д������ܷ���֬���ĵ���ϸ��������������ı�Ƥ�ܷ���һ����������к�������ϸ����desquamated cells����
���������Ƥ����������ɱ�Ƥ�������Ľ��������ë�������צ����Ƭ�ȡ�һЩ����Ĺڣ�comb�������⣨wattle����Ϊ�Ӻ�ġ�����Ѫ�ܵ���Ƥ�����ɣ����ڸ��ж������Ǻϣ�anastomosis���ṹ��
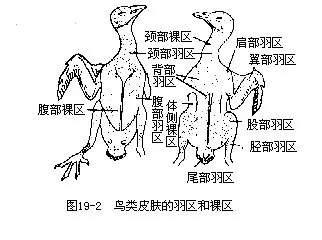
������ë�����������һ�������ڣ���Ϊ��feather tract������Щ�ط���Ϊ������pteryla������������ë�ĵط���������apteria����ͼ19-2������ë������������ʽ�������ھ��ҵķ����˶������ู����������������������й�ϵ�������ڷ����ڼ䣬������ë�������䣬��“���Ѱ�”����������ص���ж���Ұ�����ɼ��������Ƿ��ѽ��뷱ֳ�ڡ����µ���Ҫ�����ǣ��ٱ������£��γɸ��Ȳ㡣ͨ�������������Ƥ�������ɸı���ë��λ�ã��Ӷ��������£��ڹ��ɷ������ٵ�һ����——����β�𣻢�ʹ�������������ͣ����ٷ���ʱ���������ܱ���Ƥ���������ˡ���ɫ���ɳ�ΪһЩ���ࣨ����������༰��������Ѵ��ı���ɫ��
����������ë�Ĺ�����ܣ��ɷ����¼��֣�

 ��
��
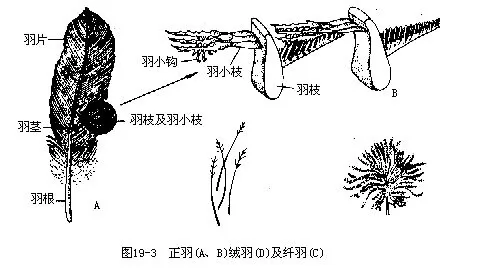
����1.����contour feather���ֳ�����Ϊ����������Ĵ�����Ƭ�����β����������һ��ǿ������𣬷ֱ��Ϊ����flight feather����β��tail feather��������β�����״����Ŀ����������������֮һ���������������Ƭ�����ɡ������¶β�����Ƭ�IJ��ֳ�Ϊ�������������Ƥ���С���Ƭ��������ϸ������֦�����ɡ���֦�����������г��ŵ���С֦����С֦��������ͻ��ڽᣬʹ���ڵ���С֦��������������ɼ�ʵ�����е��Ե���Ƭ���ԓ��������ͱ������壨ͼ21—3�������������뿪����С֦���ɽ������������й��ᡣ���ྭ����ȡβ֬�������ڵ���֬����������Ƭʱ����ͿĨ��ʹ��Ƭ������õĽṹ���ܡ�
����2.����plumule��down feather��λ�������·�������״�����������ĸ��Ȳ㡣������ˮ���ر����Ҫ���ü�ֵ��Ѽ����������ë������Ľṹ�ص���������������С֦�Ĺ�״ͻ���������ܹ��ɼ�ʵ����Ƭ���׳���������С֦��ͼ19—3����
����3.����filoplume��hair feather���ֳ�ë״��������ë��������������������֮�С��ڰε�����������֮��ɼ�����ͼ19—3��������Ļ�������Ϊ������
����������ë�DZ�Ƥϸ���������Ľ��ʻ������ϵͳ��������������Ľ�����Ƭ��ͬԴ�ģ���һ�ּ�˵��Ϊ������������������ڳ�����Ӧ�ڷ������ʽ�Ľ��������У�������Ƭ���������죬Ȼ�����ѳ�֦��������ë��
��
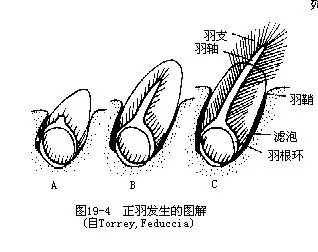
�����Ӹ��巢���ɼ�����ë���Դ������Ƥ���Ƥ�����ɵ�����ͷ����������ͷ��������������γ��������еĽ�����������Ϊδ������֦�����λ�ڱ�������������Ѹ�٣���Ϊδ����������������������������������������У���Ϊ��֦�������ǹ�����Ƭ��ͼ19—4����
�����������Ե�����ܴ����루bristle����Ϊһ�ֱ��ε���ë��������ɻ�����������֧����֧���д������ܡ�
��
�����������ë�Ƕ��ڸ����ģ���Ϊ����molt����ͨ��һ�������λ����ڷ�ֳ����������������ƶ���winter plumage�����������紺���������������summer plumage�������nuptial�������������ѧ�����������������Ǩ�㡢Խ������ֳ���̡���״�ٵĻ��������Ļ�������ʵ����ע���״���ػ��������״�٣���������������
��������β��ĸ������������ģ�ʹ��������ڲ�Ӱ�������������½��С�����Ѽ��ķ��������Ϊһ��ȫ�����䡣�����ʱ����ɥʧ�����������������˼������ĺ����ݴ��С����о���Ѽ��Ǩ��Ĺ����У����������ʱ�������������д��ģ�Ļ�־���������ڷ�ֳ�ڼ������ڵ���Ѽ�࣬Ӧ�Ͻ��IJ���
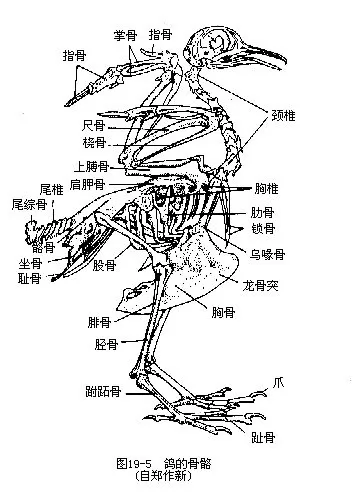
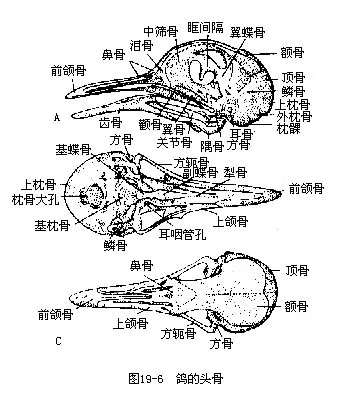
�������������� ������Ӧ�ڷ�������ڹ���ϵͳ�������������ػ�����Ҫ�����ڣ����������̣������ھ��г��������ǻ϶��pneumatization����ͷ�ǡ����������̺�֫�ǵĹǿ�����������֫��������нϴ�ı��Σ�ͼ19—5����
����1.�������ع� �����ɾ���������������β�岿����ɡ�����Ŀ����ϴ�8ö��һЩС�����ࣩ��25ö����죩���ȣ��Ҹ�Ϊ14ö����Ϊ16��17ö������֮��Ĺؽ���������Σ����찼���ǡ�����������ʽ�Ĺؽ���ʹ�Ǽ���˶�ʮ�������⣬����ĵ�һö���ʻ�״����Ϊ����ڶ�ö����Ϊ������ͷ����������������ͷ��һ��������ת������ʹ�������ͷ���Ļ��Χ������ͷ���˶���ת����Χ�ɴ�180°��èͷӥ������ת270°���������������������ԣ�����ǰ֫��Ϊ���ͼ��������ಿ�ִ������������صġ�
������5��6ö����Ӳ���ʵ��߹����ع����ᣬ�����ι̵�������������߹Dz������ǣ����ҽ蹳״ͻ�˴���������������������������ϵ���ع��Ƿ��輡�⣨�ؼ�������㣬������ʱ�������ɳ���������������ǿ���������ڱ�֤�ؼ��ľ����˶�����ɺ�������ʮ�ֱ�Ҫ�ġ������ع����ߴ��и��ʵ�����ͻ��keel�����������ؼ��Ĺ����档�ڲ��Ʒ�������ࣨ�������عDZ�ƽ��
�������ϼ��ǣ��ۼ��ǣ���synsacrum�����������еĽṹ�����������������������Լ�һ����β���϶��ɵģ��������������Ĺ��̣��Ĺǡ�������ܹǣ������ϣ�ʹ�����ڵ��沽��ʱ���֧�����صļ�ʵ֧�ܡ�����β���˻������öβ�����ϳ�һ��β�۹ǣ�pygostyle������֧�����ε�β�����༹�����������Լ�β���˻�����ʹ�������ļ��������룬�������ڷ����б���ƽ�⡣
��
����2.ͷ�� ����ͷ�ǵ�һ��ṹ�����������ƣ����磬���е�һ������������ʯ�����пɼ�ͷ�Ǻ����˫��ѵĺۼ��������ɵ�һ�Ķ������������Լ��յ������ȡ�������Ӧ�ڷ���������������ػ��Ƿdz������ģ���Ҫ�����ڣ�
������1��ͷ�DZ����ᡣ���ǿ��ķ���ڳ������������Ϊһ�����壬���ҹ����з���״������Сǻ����ͽ����������ʵ��ì�ܡ�
������2�������Ǽ���ǰ�죬������������������������м�����Ľṹ�������߽����ʣ�������������Ե���������ȡʳ���١��ִ�����������ݣ�ͨ����Ϊ��Ҳ�ǶԼ������أ������˻���ͬ�����ⲻ�������Ӧ��
������3�������Ӿ����ٵĸ߶ȷ��������������ĸı䣺ǻ�����ʹͷ�Ƕ�����Բ���Σ����Ǵ���������档�ۿ������ʹ��һ���������ڱ�ѹ�������루�������ǻ���ƣ������ɿ�������������ijЩ�����༴�Ѵ��ڣ�������������������ⷢ��Ӷ���ǿ��������ص㣨ͼ19—6����
����3.���Ǽ�֫��������Ǻ�֫��Ҳ�����ϼ�����������Ҳ�Ƕ��������ʽ����Ӧ��
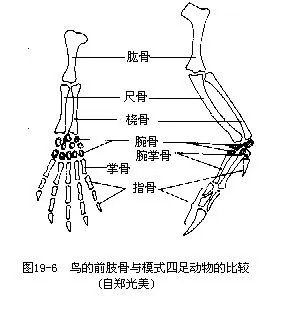
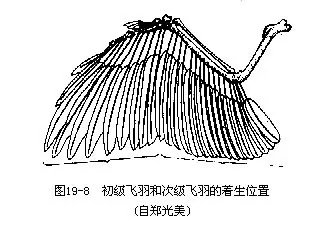
��������ɼ��ιǡ���Ǻ����ǹ��ɡ����ǵ����ᴦ���ɼ�ʣ��������Ź���ؽڡ���������������Լ��˻��ļ������ڸ����ߴ����ϳ�“V”�Σ���Ϊ��ǣ�wishbone�������������еĽṹ����Ǿ��е��ԣ����������ғ���ʱ�ɱ������Ҽ������Ҫ����ǣ���ײ��ǰ֫�ػ�Ϊ������Ҫ�������ֲ���������ǡ��ƹǺ�ָ�ǣ������Ϻ���ʧ����ʹ���Ĺ�������һ�����壬�����������������ָ���˻����ִ��������צ��ͼ19��7�����������࣬����������������Opisthocomus hoa-zin��������ָ���о�2צ��������Ե�������ֲ������ƹǼ�ָ�ǣ���������һ�з���Ƴ�������primaries�����±۲����߹ǣ���������һ�з���ƴμ�����secondaries���������Ƿ������Ҫ��ë�����ǵ���״����Ŀ���ر��dz����������������ѧ����Ҫ���ݣ�ͼ19—8����
 ��
��
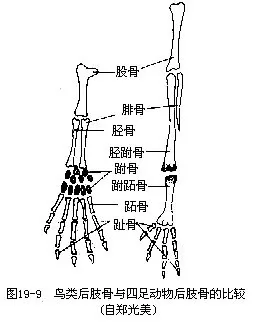
�������������ı��Σ����ú�֧֫�����غͲ����;�Ӳ�ǵ��������й�ϵ���������Ĺǡ����Ǽ��ܹǣ����ϳɱ��������ĹǼܣ����ĹDz��ֲ���ǰ����չ�������ϼ��������ϣ����ʹ��֫�õ�ǿ������֧�֡��ܹ��˻��������������ǡ��ܹDz�������½�������������ڸ����ߴ��������ᣬ����һ��������չ��������ν“����ʽ����”���������������Ӳ���������й�ϵ�ġ�Ȼ���ڼ�����½��ԭʼ���ࣨ�����������ҳܹǻ������ڸ����ߴ�����������������ĺ�֫ǿ�����ɹ����������ž���ؽڡ����ȹ����нϴ�仯������˻��ɴ�״���൱��һ�����㶯����ֹǣ��������ڵ�һ���˻����ƹ������ϣ�����һϸ���ε��ȹǣ���Ϊ���ƹǣ�tibiotarsus����Զ��һ�ŵ��˻��ƹ��������ڵ��Ź������ϣ�����һ��ϸ���ε���ǣ���Ϊ���Źǣ�tarsometatar-sus�������ּɵ�һ�ģ����ƹǼ����Źǣ��ǿ�ؽ��Լ�������������ӳ�����������ɺͽ���ʱ�ĵ��ԡ�������������4ֺ��Ĵֺ�������Ӧ��������֦��ͼ19—9������ֺ����Ŀ����̬�������������ѧ�����ݡ�
�������ģ����� ����ļ���ϵͳ������������һ�������ɹ����������Ƽ��������༡��ƽ���������ļ���ɡ�����������Ӧ�ڷ�������ڹ���������̬�ṹ���������ı䣬��Щ�ı���Ҫ�ɹ��Ϊ��
��
 ��
��
����1.�������Ժ�ļ��������ϣ������±���������˻���������������Ӧ���
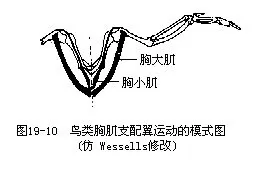
����2.ʹ��������С�����������ش��ļ���ʮ�ַ��ռ�������ص�1/5�������ǵ������������ع��ϣ�ͨ����������᷽ʽ��ʹ�퓁����ͼ19—10�������⣬������֧��ǰ֫����֫�˶��ļ��⣬�伡�岿�־���������������IJ��֣��������쳤�ļ�����“Զ����”����֫���˶�����Ա������ĵ��ȶ���ά���ڷ����е�ƽ�⣬������Ҫ���塣
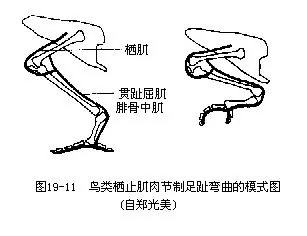
����3.��֫����������������֦�ļ��⡣��Щ�������йصļ��⣨�����ܼ�����ֺ����������м������ܹ��輡�졢������������ؽ������������ϣ���ʹ������ֹ����֦��ʱ���������ص�ѹ�Ⱥ��ȹǹؽڵ���������������ֺ�йص���������ļ�����������ֺ��Ȼ����֮����������ץס��֦��ͼ19—11�����ܼ���ambiens���������������У���ʼ���������࣬�ڸߵ����ࣨ��������Ŀ��ȸ��Ŀ����ʧ��
��
��
����4.������������ܼ��⣬��֧�����ܣ��Լ���Ĥ���ı���״����������������������������ȸ��Ŀ���ࣨ���ݣ��ر
�����������ǰ��֫��������������Ϊ�о��������ѧ�����ݡ����������йؼ���ĺ�֫��Ⱥ���^���������Ÿ��ijἡ�ȷ�����Լ������Ĺ�����̬ѧ���������˽�������о���
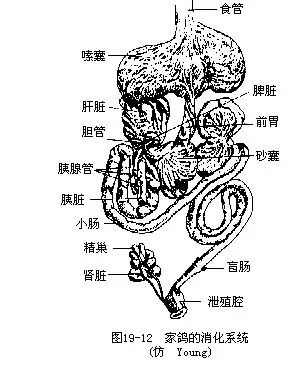
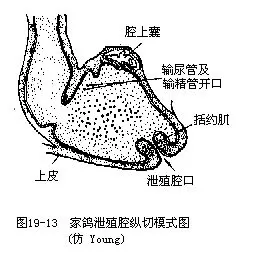
�������壩������������ϵͳ����Ҫ�ص��ǣ����н�����Լ���Ӧ�������Ǻ;���Ⱥ�����������˻�������ʳ��ʽ��ʳ����������������йء����״��ʳ�Ժ����ʽ��ͬ���кܴ���졣������������������н������ʣ������̬�ͽṹ��ʳ�Ժ����ʽ�йأ�ȡʳ�������������ʱ������״��ˢ״����ľ�����ߵ������ܰ���Ƥ�µĺ��湳����ijЩ��ľ��ͷ�����࣬������Ĺ���������������Զ����߿ɴ��峤��2/3����ǻ������Һ�٣�����Ҫ��������ճҺ������ʳ�ȵ���ȸ����Һ���ں�������ø����������������Ŀ����Һ�������ں���ճ���ǵ��ף�glycoprotein������������Һ������ճ�϶��쳲�����еĽ�˿�������ij�����Ϊ��ͳ���̲�Ʒ“����”��Ŀǰ������Ϊ������˿�࣬�ѽ�ֹ�ɼ�����Щ�����ʳ��һ�����ػ�Ϊ��ң�crop�������������غ�����ʳ��Ĺ��ܡ��Ƹ��ڷ�ֳ�ڼ䣬��ұ��ܷ���һ��Һ�壬��Ϊ“����”������ι�dz��롣ʳ�����ࣨ����˺����ɣ���������Ƴɵ�ʳ���dz��������θ��Ϊ��θ��ǰθ����glandular stomach�� proventriculus���ͼ�θ��ɰ�ң���muscular stomach�� gizzard�������֡���θ���ڸ������壬�ɷ���ճҺ��Ϊһ��ǿ�ᣩ������Һ����θ���Ϊǿ��ļ���㣬�ڱ�Ϊ��Ӳ�ĸ��ʲ㣨��ҩ“���ڽ�”����������֣���ǻ�ڲ������������ʳ��ɰ�����ڼ���������£����ʱ���ɰ��һ��ʳ��ĥ�顣ɰ���������ӵ����������й�ϵ��ʵ��֤����θ������ɰ���ļ�������������������3������һ����P���ӵ������������10������ʳ������ļ�θ����������ֱ�����̣��������㣬�Ҿ�������ˮ�ֵ����ã������ڼ���ʧˮ�Լ�����ʱ�ĸ��ɡ���С������紦������һ��ä��������ֲ����άΪ��ʳ�����ࣨ�缦�ࣩ�رä��������ˮ���ã�������ϸ��һ�������ֲڵ�ֲ����ά��������Ϊä��Һ�������ļ���ά����B�����á����ſ�����йֳǻ����һ�㻹���������������������ͼ19—12��������йֳǻ�ı�����һ����������壬��Ϊǻ���ң�bursa fabricii����ǻ�����������������ʧȥ��ǻ��Ϊһ�������ܰ���Ƥ������ṹ��ͼ19—13����ǻ���Ҿ����ѱ�������һ���ܰ���֯��������������������ƺ��ܲ����������߳ɷֵķ�������к�������������Ƥ�ʼ��ػ��״�ټ��صĻ��ԡ�ǻ���һ��������������������һ��ָ�꣬�ر��ڼ�������Ŀ��������䷽���ѱ��㷺Ӧ�á�
����������������������ص���������ǿ����������ʮ��Ѹ�٣�����������ǿ���³´�л��ʢ�����ʻ�����ʵ��֤ʵ���Թ����ʵ����������ι��ȸ��Ŀ���࣬��1.5Сʱ��ͨ���������������Ѽ��Anas rubripes����ʳ�ᆳ30���Ӻ��ų����߶ȵ����������������ģ�ʹ����ʳ����ʳƵ����ȸ��Ŀ����һ�����Ե�ʳ��Լ�൱���ص�10����30��������һ�����Ե��۽����������ص�һ��������1500g��ȸӥ������һ��ҹ�Ե�800g��1000g�⡣��Щ�������������������ء��ݼ���������Calypte anna����Ϣʱ��ÿСʱÿ����������10.7mm3��16.0mm3�����������ڷ���ʱ������85mm3��
����������Ҫ���������Ǹ�������࣬���Ƿֱ���ڵ�֭����Һע��ʮ��ָ�����ڹ�����������������û�б��ʵ�����
�������������� ����ĺ���ϵͳʮ���ػ��������ھ��зdz���������ң�air sac��ϵͳ���������ͨ�������ҹ㲼�����ࡢ��ǻ�Լ�ijЩ�˶�����֮�䡣���ҵĴ��ڣ�ʹ����������صĺ�����ʽ——˫�غ�����dual respiration������������½�ܼ������������ʱ����������������ͬ���������ϵͳ������ṹ�����������������ĸ�����������Ӧ�ģ�ʵ�������һ֧�����е����������ĵ�����������Ϣʱ��21��������Ҳ�DZ�֤�����ڷ���ʱ��Ӧ�㹻������װ�á���������ֹʱ����Ҫ���عǺ��߹��˶����ı���ǻ�ݻ�������κ����ҵ��������С������������л��������ʱ���ع���Ϊ���ἡ�⣨�ش���С��������㣬�����ȶ��������Ҫ�����ҵ�������Э������ɺ���������ʱ�������ţ��������ζ����룻����ʱ����ѹ���������ٴξ����ζ��ų�������������Խ�죬����Խ���ң����彻��ҲԽ�죬������ȷ���������ij�ֹ�Ӧ��
��
��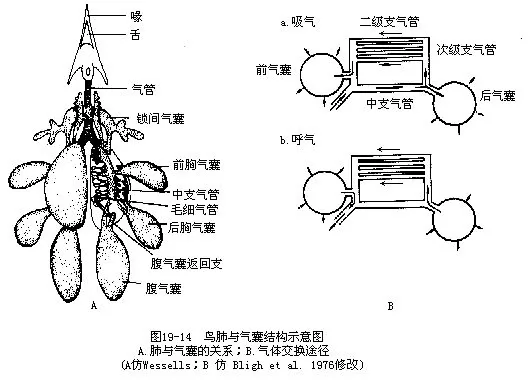
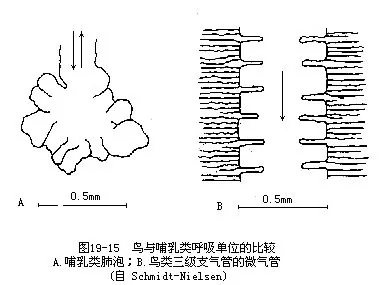
��������������ҵĹ���ʮ�ָ��ӣ�����ֻ���ز����ṹ���ص�ͻ��ܣ�ͼ19—14��������ķ��������ǽ�С�ģ���һ�ֺ���״ȱ�����ԵĽṹ�����ֽṹ��Ҫ���ɴ�����ϸ֧������ɣ�������ϸ�ķ�֧��һ�ֳ�ƽ�����е�֧���ܣ���Ϊ����֧���ܻ�ƽ��֧���ܡ�������֧������Χ�з���״���е����ܣ�����ֲ����ڶ��ëϸѪ�ܣ����彻�����ڴ˴����У�������εĹ��ܵ�λ�������������˵���൱������½�ܼ�����ر��Dz����ࣩ�ķ��ݣ����ڽṹ�����б��ʵ����𣬼�������ϵϸ֧����ĩ������ä�ң������������ȴ�뱳�༰����Ľϴ�֧������ͨ�����������ä�ˣ�ͼ19��15��ͼ19��16�������������ֱ������3μm��10μm����ε����彻���������cm2/g���أ�����Լ��10����
�����������֮��Ϊ�ᴩ�������֧���ܣ�Ҳ�г���֧���ܣ�����֧�������������ܶ��֧���Ʊ�֧�����븹֧���ܣ��������ܳ�Ϊ�μ�֧���ܡ�������֧���ܽ���Ŀ�ڶ��ƽ��֧���ܣ�����֧���ܣ�����ᣬ�����ڷ�����һ���������������ӱ�֧����→ƽ��֧����→��֧���ܣ���Ϊ“d-p-vϵͳ”��Ҳ���Ǻ���������ʱ�������ڷ��ھ�Ϊ����������a uniderectional path-way����
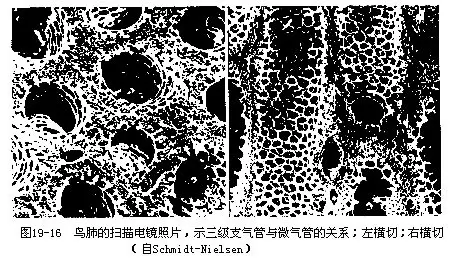
��������������ĸ�������ϵͳ����Ҫ�ɵ�����״��Ƥϸ�����ɣ������������֯��Ѫ�ܣ���ȱ�����彻���Ĺ��ܡ�����һ����9�����ң���������֧����ĩ����ͨ����Ϊ�����ң������Ҽ��������ң����븹֧������ͨ����Ϊǰ���ң������ҡ��������Һ�ǰ�����ң�������������Ϊ������֮�⣬��ϵ���ҳɶԡ����ұ鲼���������١��ؼ�֮�䣬���з�֧�����Ĺ�ǻ�ڡ�
����������ԣ�����������ʱ�����ʿ�������֧���ܴ�ֱ�ӽ�������ң�һЩ����“�·�”�����࣬��һС�����徭���·ε�����֧���ܺ��ٽ�������ң������ͬʱ��һ�������徭�μ�֧���ܣ���֧���ܣ�������֧���ܡ��ڷΣ�Ҳ��“�ŷ�”�������ܴ�����̼������������ʱǰ��������ͬʱ���ţ�����ʱͬʱѹ�������������ʱ�����ں�CO2������徭��ǰ�������ų�����ʱ��������������������徭��“����֧”������ڽ������彻�����پ�ǰ���ҡ����ܶ��ų���ͨ���Ա��������ʵ�鷢�֣�һ������Ŀ���Ҫ����2�κ����˶�������ų����⡣��Ȼ��Ϊ����������������̣�����ÿһ�������������������������������ϵĸ�������������ͨ�������������������ﲻͬ�ġ�
��
���������������ǰ���ҵ���������������Э���ģ����ʹ�����ھ��ҷ���ʱ��ǰ���������œ�����ɶ����������缸�������������ϵذѿ���������������ų��������һЩ�о�ָ���������ڷ���ʱ���䓁���Ƶ�ʲ���һ�������Ƶ����Э���������ѻ�����������Ƶ��Ϊ1��1������Щ���������ࣨ������ɽ�Ⱥ�ҰѼ����Ϊ5��1������Э����ϵ�ڷ��������б仯�ġ�
�������ҳ��˸����������⣬�������ڼ�������ı��أ����ټ�����Լ�������ĥ������Ϊ�����ȴ�л����ȴϵͳ�����˼���һֻ���ŵĸ룬���������3/4��������ȴ����
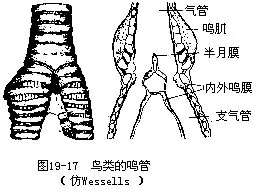
�������ܣ�syrinx�������������ػ��ķ������٣�λ��������֧���ܵĽ��紦��ͼ19-17�����˴��������ܱھ��䱡����Ϊ��Ĥ����Ĥ����������������������ಢ���������������������ɵ������ܱ���״�����ų̶ȷ����ı䡣���ݣ�ȸ��Ŀ���ࣩ�����ܼ������������ӣ���������˫�غ������ص㣬ʹ����������ʱ���������ܶ������ö�������������һ������������Ҳ������ͬ��һ��½�ܼ�������粸���ࣩ�ķ������پ�λ�������϶ˣ��Ҿ���������ں���ʱ����������ĺ�����4�鲿�ֹǻ������ǹ��ɣ���Ƿ������٣�����ͨ�����ŵ��˶�������������
�������ߣ�ѭ�������ѭ��ϵͳ��ӳ�˽ϸߵĴ�лˮƽ����Ҫ�����ڣ�������ѪҺ��ȫ�ֿ�����ȫ��˫ѭ����������ǻ�������嶯����������������������Ƶ�ʿ졢����ѹ�ߡ�ѪҺѭ��Ѹ�١�������塢Ӫ�����ʼ�����Ĵ�л��ʢ��
����1.���������������Դ�Сռ�������е���λ��ԼΪ���ص�0.4����1.5�����ķ�����������ȫ�ָ��������ķ����������Լ����ķ��������ң����͵ȼ���������ľ�������������ȫ��ʧ�������徲����ѪҺ�������ķ��������Ҷ��ɷζ�����Ρ��ڷ��ھ������彻���������ḻ��ѪҺ���ξ������Ķ�ע�����ķ����پ������Ҷ������嶯����ȫ������������ķ��������Ҽ�İ�ĤΪ�����ʹ��ɣ��˵�������½�ܼ����ﲻͬ�����������������ƽṹ����
��
��������������Ƶ�ʱȲ������ö࣬һ�����300��500��/min֮�䣨��19—1��������ѹ�ϸߣ������ۼ�Ϊ25kPa���Ƽ�Ϊ22kPa�������ѪҺ��ͨѸ�١�
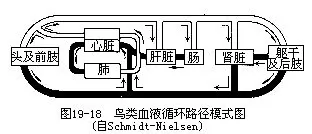
����2.��������Ķ���ϵͳ�����ϼ̳��˽ϸߵȵ����ж�����ص㣬������嶯������ʧ�����Ҳ��嶯�����������ҷ�����ѪҺ���͵�ȫ����ͼ19—18����
��
����3.��������ľ���ϵͳҲ�����������������ƣ�����2���ص㣺
��
������1�����ž��������˻�����β������ѪҺֻ������������������ϵ����������ġ�������˱��棬���������ž���ǻ�ھ���һ�ֶ��صĺ���ƽ�����İ�Ĥ���ɸ�����Ҫ���Ѿ���ѪҺ����������ƹ����ࡣ
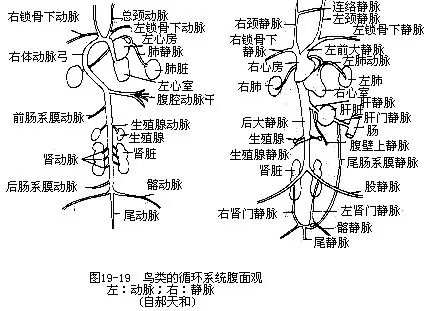
������2����β��ϵĤ���������ռ�����ѪҺ������ž�����β��ϵĤ����Ϊ���������У�ͼ19—19����
��
����4.ѪҺ���ܰ�����ѪҺ�еĺ�Ѫϸ�������ϲ������٣�ԼΪ2000000��7645000��/cm3������Ѫϸ���ߺˣ�һ��Ϊ��Բ�Ρ���Ѫϸ���к��м�������Ѫ�쵰�ף�ִ����������������̼�Ļ��ܡ�
�����������һ�Դ���ص��ܣ��ռ�������ܰ�Һ��Ȼ��ע��ǰ���������������С����ë�в��߲������������ӹܣ�����������ࡢ�����ʺ�֬���Ĵ�л������������ž���ֱ�ӽ����������غ����á������ܰ�ϵͳ�ϵ��ܰͽڣ�����ֻ��������Ⱥ�������о���Ѽ���Ƕ�����Ÿ����������Ϊ���Dz�����������ܰͽ����������ܰ͡��������ࣨ����������Ѽ�ࣩ����������ܲ������ܰ��ġ����ߵ����ľ�������Ҳ��һ���ܰ��ģ���������ʧ��
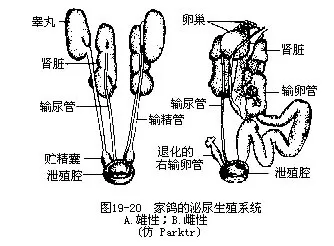
�������ˣ���й�������������������ƣ���̥��Ϊ������������ʹ�����ܵ�Ϊ�������������������Ȳ������ռ���ص�2�����ϡ���С�����Ŀ�Ȳ������2�������������ʢ���³´�л�����У���Ѹ���ų����������ˮƽ���������ġ����ྭ����ܿ�����йֳǻ��ͼ19—20����
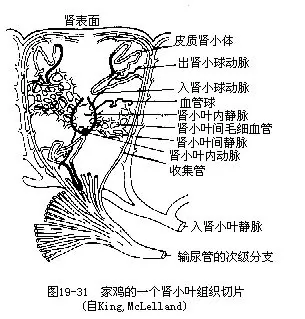
�������������ͨ����ͷ���С�β3����Ҷ��ɣ����ҳɶԡ�ÿһ��Ҷ�����ڶ�ġ���۳����ε���СҶ������СҶ��������ž�����������С������������ռ��ܡ���СҶ���������뾲�������ܵ�С������ëϸѪ����ͨ������СҶ����λ�����뾲�����������֧�γ���С��ͳ���С������ͼ19—21�������ֽṹ�벸���ͬ����ʾ����û��ͬԴ��ϵ��
��
��
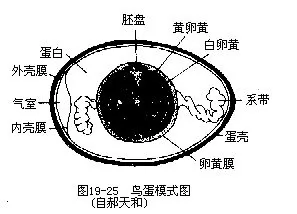
�������������������������ṹ�ɵģ������Dz���������ء��������������������ˮ�����ʰ����̵İ�ɫ�ᾧ���������̥���ѿ��ڷ������в����ų�����ͼ���ˮ�ֵ�ɢʧ�������ġ��ټ��ϳ������С�ܺ�йֳǻ������������ˮ�ֵĹ��ܣ�������������ʧˮ���١�����߰��ף�����������ͬ�����ʱ�ų����⣬ͨ����Ϊ��Ҳ�Ǽ������ص�һ����Ӧ����ָ�����ǣ��������˲���ƫ�������͵��������Զ�������ĸ�����з�������Ϊ������ж��ֳɷ֣����Ҵ�X���߷������������ֲ��������ṹ�ɣ��������йֳǻ��İ�ɫ���ʵĻ�ѧ�ɷ־�����ʲô�����д���һ���о������������ˮƽ����ڷ��棬�����ᵽ���������е����١�����λ���ۿ��ϲ��������ڱǼ�������ܷ��ڳ������Ũ�ȴ�ö���Ȼ��ƣ��������к���5������Һ�������ѽ������ڵĺ�ˮ���������η��ų���ά����������ѹ��һЩɳĮ����������ࣨ���������Լ�����Ŀ�����࣬������Ҳ�е�����ѹ�Ĺ��ܣ�ʹ֮����ȱ����ˮ������ʧˮ�ϸ��Լ�ʳ�����ηָߵ����������档
����������һ����Ĥ����һ���������ű�������ˮ�ֵ����⡣��������Ƥ�����ȱ�����壬������н�����ë����Ƭ����Щ�����������ˮ�ֵ����������������ŷ�����ʧˮ�ֺ��٣����ˮ��������������½������Ϊ�١����Dz����������Ϊ�����ƾʳ���е�ˮ�������������Ҫ������������ʵ���Լ����ǶԲ��͡���������β����������ʵ��֤������ˮ�dzɻ�Ĺؼ�֮һ������Ҫ�����ں�����������ˮ�����Լ�����ʱˮ��������ȴ���á����루Zenaida macroura����ʵ�鷢�֣���39�������£�ÿ�����ˮ��Ϊ��23��ʱ��4�����ڸ�����24Сʱ����ˮ��ʹ���ؼ���15������һ����ˮ��������֮�ھͿɻָ����ء����ֶ�ˮ��Ҫ���ڻ�Į������������Ϊ���ԣ����dz������ɴ�ȺǨ����Ѱ��ˮԴ���ҹ��Ͷ������Թž۲쵽ë��ɳ����Syrrhaptesparadoxus����������Ǩ�������֮�����ϵ����������ʮ��ǰ����������̬ѧ�������á�
�������ţ���ϵͳ�й�
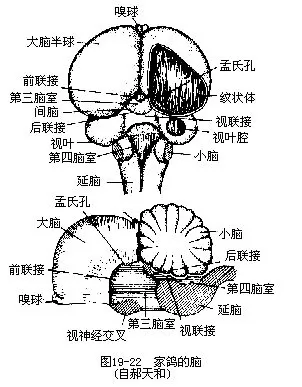
����1.�Լ�������������ںܶ���������࣬���벸�����кܴ�ͬ�����磬�������м�����Ĵ���Ƥ�㣨cerebral cortex����������������Ժ�С�Եı��涼�Ƚ�ƽ���������������������ޣ��Թ����Իأ����������Ҷ�˻������ԵĶ��ںܱ������ײ�ʮ�ַ����Ϊ��״�壨striatum corpora������״�������ิ�ӵı��ܻ��“ѧϰ”�����ࡣ����ļ����������ԡ����Ժ������²������ֹ��ɣ����������²���Ҳ�������𣩣�hypothalamus�����ɼ��Եĵױڣ�Ϊ���µ������ಢ����ֲ������ϵͳ�������²��������´���ķ������Źؼ��Ե�Ӱ�죬ͨ�����´���ķ��ڶ����������ڷ����١���������Խ��������Ӿ��Լ�һЩ�ͼ����ഫ��ij嶯�����ɱȽϷ������Ҷ��С�Ա��������ö࣬Ϊ�˶���Э����ƽ�����ࣨͼ 19��22����
���������� 12���������� 11�ԣ���������������Ҷ���һ�����Ƿ�����������飬ֱ��1965��ʼ֤������ڡ�
����2.�й�����ĸй������Ӿ���Ϊ���������֮�������Ϊ�˻�����Щ�ص������������������ϵ�ġ��Ӿ�Ϊ���趨�����Ҫ���١�
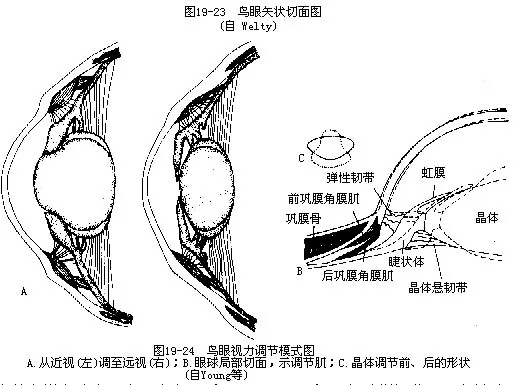
�������۵���Դ�С�����м����ﶼ�������۳ʱ�Բ�Σ�Ϊ��ƽ�ۣ�flat eye����ӥ������Ϊ��״��globular eye�����^ΪͲ״��tubular eye�������������Ϊ���͵Ĺ�Ĥ����ǰ����������һȦ����״���еĻ��ι�Ƭ���ƹ�Ĥ�ǣ�sclerotic ring������������ڵļ�ǿ֧�ܣ�ʹ�ڷ���ʱ������ǿ������ѹ����ʹ������Ρ��ں��۷��ڵ�����������һ������ɫ�صġ����ĺ���Ѫ�ܵĽṹ����Ϊ��Ĥ��pecten������Ĥ��ȷ�й��ܻ���������������ݻ��������������ڵ�Բ��ͻ��conus papillaris��ͬԴ��һ����Ϊ��Ӫ������Ĥ�Ĺ��ܣ����ɽ�����ĸı�����������ڵ�ѹ����Ҳ��һЩ֤��ָ�����������ڹ�����Ӱ�������չ���ɵ�Ŀѣ��ͼ19—23����
��
�������۵ľ�����ڼ���Ϊ���Ƽ����˵����������������������������ﲻͬ�����ڷ�����Ѹ�پ۽��������������ġ������ǰ��Ĥ��Ĥ����anterior sclerocorneal muscle���ܸı��Ĥ�����ȣ���Ĥ��Ĥ����posterior sclerocorneal muscle���ܸı侧������ȣ��˵����Ʋ������½�ض��������������ܸı侧�����״���Լ��������Ĥ��ľ��룩�����һ��ܸı��Ĥ�����ȣ���֮Ϊ˫�ص��ڣ�ͼ19—24��������ĺ�ʳʻơ��ּ���ɫ����ʼ�Ҳ�Ǻ��Ƽ����벸���ͬ��DZˮ������ˮ��ʱ���ܽ��ʼ���������ѹ�Ⱦ���ǰ����Э��������
������������������־��ɶ�Ѹ�ٵĵ��ڻ��ƣ�ʹ������һ˲��ѱ�ƽ��“Զ����”����Ϊ“������”��ӥ������������ɱ����ڳ�Ͳ״�����Ƿ����������ز����ٵ�������ӥ�ڸ߿����ܲ������ڵ����࣬���ڼ������ڸ���ץ�������������˴�8���������ڼ����������ɳ棬�ⶼ��������õ��������ڷֲ����ġ�����Խ�ٽ�Ŀ����ͻ�Խ���������
���������������ٻ������������࣬���е�һ�����ǣ������ǣ��ͳ��ε��������ҹ�������࣬����������Ϊ���

��
 ��
��
��������������ǻ�ھ��� 3���Ǽף�nasal concha����������˻���һ����Ϊ��Ҳ�Ƿ�������IJ���������ࣨ����أ�գ�����൱����ѳ�Ϊһ�����Ѱʳ�Ķ�λ���١�
������ʮ����ֳ������ֳ�ٵĻ�����������Եļ����Ա仯���ڷ�ֳ����ֳ�ٵ���������ٱ�����ǧ����һ����Ϊ��Ҳ����Ӧ�ڷ��������йء�
����1.������ֳϵͳ�����������������ƣ����гɶԵ�غ����侫�ܣ��侫�ܿ�����йֳǻ���������Ѽ��ȵ�йֳǻ����¡�𣬹��ɿ����йֳǻ��Ľ��������������;��ӵ����á���ijЩ����Ŀ������Ŀ�����࣬���д��Ž������ĺۼ�����Щ��������Ϊ��������Ա�ı�־�����ڴ�������࣬�����߽������٣���������йֳǻ�ڽӺ϶��ھ�������ľ�����йֳǻ�����ѹ��ڴ�������Ȳ���������罫�Ƽ�Ѽ����Ѽ����֮��һ�ܲ�64���ܾ��ѣ���3��Ϊ3�������һö�ܾ����ڵ�17�������
����2.������ֳϵͳ�������������ߵ�һ�ģ���ࣩ�й��ܵ��ѳ����Ҳ��ѳ��˻�����ijЩӥ�ࣨ������ȸӥ���κ����������а���������гɶԵ��ѳ���һ���ѳ��˻���ͨ����Ϊ��������;�Ӳ�ǵ����йء�������ͨ�����ѹ�ǰ�˵����ȿڽ������ѹܡ��ܾ����÷��������ѹܵ��϶ˡ����ܾ��������ѹ������еĹ����У����α����ѹܱ������ڵĵ��ף�albumen������Ĥ��shellmembrane�����ѿǣ�shell�����������������ѹ����ƶ�ʱ�����ڹܱڼ�����䶯����ת���������Ծ��ȵĵ��ײ㣬���˳����ײ�����Ťת����ϵ����ͼ 19—25������ϵ���������ŵ��ѻƣ�����������ϵ��ʹ������Զ���ϣ������ڷ���������һ����Ҫ������ѧ��Ӧ���ѿ�Ϊ̼��ƣ�89����97����������������л��ﹹ�ɣ����������ǧ��С�ף��Ա�֤���ڷ���ʱ�����彻�����ܶ�������ѿ����и�����ɫ�ͻ��ƣ����������ѹ����¶˹ܱڵ�ɫ��ϸ���ڲ���ǰ5Сʱ���������ڵġ������йֳǻ�ų���
��
������������ѹ�Ϊ����״�������ѵ����ѹ���Ҳή���������¶˵�ֱ�����ȡ�����ص����ΪҰ���ʱ������������ݡ�
���������ܴ̼�������������Լ����ﶬ���ڲ�����ĵ�����֪һЩҰ���绷�������Ƹ���������ȸҲ�Թ��մ̼��з�Ӧ������һЩ���������С���������ܴٽ�“����”������˶��ͽ�ʳ�����ͨ�����´�����ڼ��ش̼��ѳ���12��14Сʱ���ն����´�����ںͲ��ѵĴ̼����������Ϊ���Ĵ̼����ڰ⡣����ָ���������ߣ��������Ŀɼ��⣩�����10����19�������ʣ�������ά����D�ĺϳɡ�
����������з��ѡ�������һϵ�б��ܣ���֤�˺���нϸߵijɻ��ʡ�
��
�ڶ��� ��ٷ���
��
�����ֽ���֪����Լ 9 000���֣���Ϊ�����Ǹ٣��������Ǹ٣�Archaeornithes���ͽ����Ǹ٣�Neor-nithes����
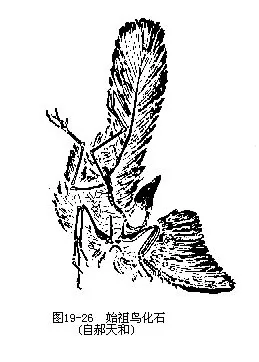
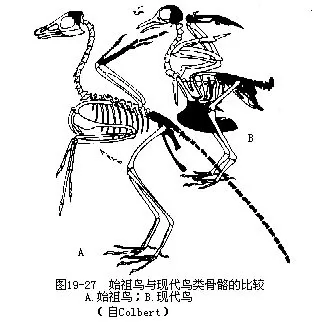
���������Ǹ���ʼ����Archaeopteryx lithographica��Ϊ���������ھ�� 1.45����ǰ����٪�͵ز��У������ѱ����Ļ�ʯ��7���������ڵ¹��ͷ�����ʡ�����Ҹ�����ӡ��ʯʯ�����ڣ���1861����Ϊһ������ë����1861�������Ǽܣ�Ϊ���ֵ�ģʽ�걾����1877�������Ǽܣ���1956��1970��ϵ1855���������1973��ϵ1951���������1988�ꡣ1984�������ҹ��������������Ĺ���ʯ���ǵ¹�������״μ�¼��ʼ������������������Ĺ�����̬�������������Ƶ������ǣ��پ���ë�����������۹���Ϊ“����ʽ”���ܺ�֫��4ֺ����ǰһ�����־����������������������Ҫ�У��پ߲����ݣ���˫�����壻����18��21ö�����β�ǣ���ǰ֫��3ö������ƹǣ�ָ�˾�צ������������δ���ϣ����ع�������ͻ�����߹���״ͻ��ͼ19—26��ͼ19—27����������ʼ���������У�����“��ë”֮�⣬���������ڹ��������һЩ��Ա���ҵ������������Ϊ“����ë�����ж���”������������ʱ����“�������������������”�����ۣ� 1985�������������“ʼ����ʯ�걾�ϵ���ë��α���”���ʡ�����������ѧ�Ҳ��ö����ִ������ֶζԱ걾�ļ������ر���1988���ٴη��־���������ëӡ����ʼ����ʯ�걾����ʹ��һ���۸�һ���䡣
��
����1991��ӡ��ѧ�߱������ڱ����������͵ز��з��ֵ����ʯ������Ϊԭ��Protoavis����������һЩ��ʼ����ԭʼ�ġ����ƿ�����������
������ʼ����ȹ���ʯ���о�����ϵ��������Դ����һ������������⡣�Ƚ����еĹ۵��������Ǵӽ��Ƽ����ࣨPseudosuchia���������ࣨOrnithichia���IJ۳���һ֧���������� 20����70���������������Ϊʼ��������ſ�����Theropoda����ǻ������Coelurosauria���Ĺ۵���ռ�Ϸ硣������һѧ˵�����ܵ�����ķ��ԣ���Ϊ���۴ӻ�ʯ�ṹ�Լ�����ĵ��������˵�����������������ʡ�
���������Ǹٰ������Ѽ�������һЩ��ʯ�����Լ��ִ����ࡣ��ʯ�����Իƻ���Ŀ��Hesperor-nithiformes��������Ŀ��Ichthyornithiformes��Ϊ���������ǵĹ��������ִ����൫�ϡ����߲����ݡ��ҹ����귢���˴�������Ѽ����ʯ������������Ŀ�����е��й���Sinornis santensis����������ʡ����������������֪�������ɵ�������Gansus yumenensis�����ڸ���ʡ���ţ��Ǻ�����ˮ���ԭʼ��Ⱥ��������Cathayornis yandica��Ҳ�������������dz�ʼ����ԭ���������ԭʼ�����ʯ��
�����ִ�����Ǹ�����ɹ�Ϊ3����Ŀ������
��
һ��ƽ����Ŀ��Ratitae��
��
����Ϊ�ִ������������ࣨ���ش��ߴ�135kg�����2.5m�������ڱ����������һϵ��ԭʼ���������˻����عDz�������ͻ�𣬲���β�۹Ǽ�β֬�٣���ë���ȷֲ���������������֮�֣�����֦������С����������γ���Ƭ��������߷���Ľ������٣���ֺ��Ӧ������������ڼ��٣�2��3ֺ�����ֲ������ϰ����ޡ����Ͱ����ϲ�����
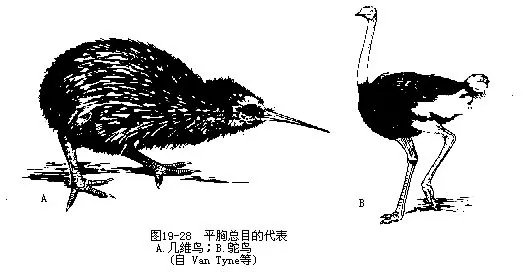
����ƽ����Ŀ����������Ϊ����Struthio camelus����Ʒ�������ͼ 19—26������Ӧ��ɳĮ��ԭ�����һ���СȺ��40��50ֻ���������Ѹ�١���ʱ�Գᓁ��������һ���ɴ�8m��ÿСʱ����60km��Ϊ������������ʳֲ����������Ӽ�С���������ɫ������ɫ�ڡ�ƽ����˵��“��������”��˵“������Σ��ʱ��ͷ����ɳ����”���������о�֤ʵ������һ�ִ����˵���������ڷ�ֳ��Ϊһ�۶�ƣ�����ѵ�����һ��������Ѩ�ڣ�ÿѨ����10��30ö�������ɫ����Լ1300g��������Ϊ6�ܣ����������ʱ���ѣ�ҹ�����������Ρ�
��������Ŀ����������������������Rhea americana������ţ���ư�������Dromaus nova-chollandeae�������������������м�ά��Apteryx oweni����Ϊ�����ڴ����������ϵ�ϡ�����ࡣ����缦������β���˻�������䣬�ǿ�λ��ļ�ˣ��˵����ڲ�ͬ����ҹ����ȡ����Ϊʳ�������������Ķ�Ѩ�����������ء������������������������“kiwi…”����������ά����1�� 2ö����ɫ�����ڶ��ڣ��ѵ���Դ�СΪ����֮�ڣ�����ռ���ص�1/4����������ѣ�ͼ19—28����
��
��
���������Ŀ��Impennes��
��
����DZˮ������С��������࣬����һϵ����ӦDZˮ�����������ǰ֫��״�����ڻ�ˮ������Ƭ״��ë������̶�������Ƭ��խ�������ȷֲ��������β�̡��ȶ̶����������ֺ����룬��Ӧ��Ӿ�����½������ʱ�������ֱ��������ҡ�ڡ�Ƥ��֬������������ں��������ˮ�б������¡��������ض����������عǾ��з��������ͻ������ǰ֫��ˮ�йء���Ӿ���٣����˳�Ϊ“ˮ�·���”���ֲ������ϰ���
���������Ŀ�Ĵ���Ϊ����죨Aptenodytes patagonicus����ͼ 19��29�����ֲ����ϼ���Ե�����������뵽��½����km������ǧ��ֻ��Ⱥ��ֳ����ֳ�Ժ���غ������������ϲ���ÿ��һ�ѣ����ڱ��ϡ�������Լ56�졣�����������Σ����������Ϊ���۹�ͬ���ѣ�����ʱ���������ڽ��ϣ������¸������µĴ�״Ƥ�����渲�ǡ���췱ֳ����������ϼ��ĺ�ҹ�����½��еģ������絽��ǰ�����Ѳ��������������ʱ�������ѳ�;���浽�������2���º��ٷ��ء������ڴ��ڼ�ȫ����ʳ��������֬��ά�֣���ߵľ�ʳ��¼Ϊ4���£�����һ���Dz������������ġ��������������ϼ���ԭ����Զ�ļ��������䲽�б���������ʱ�ɽ��������أ�˫����ٻ�ѩ����֫�ƻ�����Ŀ�ţ��������١�

��
���������ϼ�������������������ҹ��������ꡢһ���ʵ�ããѩ����ȱ��ɽ��һ��ĵ�ò��־���Լ������ص㣨�߶��ܼ���ֳ�Լ�ʧȥ������������½�����ڲ������������ʮ����һֱ�����źܶ�����ѧ�ң���������Ϊ�о�������Ϊ�Ķ������磬�ں�ҹ�����£�����������������ʶ��ģ���������ʲô��λ���ٵ��Դ�������������ò������ˮ½����ԭ����������10�����ϵ�ʱ��������һ�����ķ�ֳ������973�ι۲죬�����о��������˱�־����������� 82������ڶ��껹��ԭ��ż��������һ����һ�� 11�ꡣ�й�����Ǩ�㶨����Ƶļ�˵���еľ��Ǹ��ݶ����ʵ��Ľ�����Գ�ʵ�ġ�
����������Ҫʳ������Ϻ����������ȣ��ڼ��غ�����̬ϵͳ��������ת��ռ��Ҫ��λ�������ų��ķ�㣬�Ǽ���̦�����µȵ���Ҫ������Դ���������γɷ�������Ҫ���á�
��
����ͻ����Ŀ��Carinatae��
��
����ͻ����Ŀ�����ִ�����ľ���������ֲ��鼰ȫ���ܼ�Լ35��Ŀ��8500�����ϡ����ǹ�ͬ�������ǣ���������ڷ��裬�عǾ�����ͻ�����4��6öβ�����ϳ�һ��β�۹ǡ��߳����Թ��������������Ƭ�����������������֮�֡����������������߽������١�
�����ҹ�����ͻ����Ŀ���࣬����26Ŀ81�ơ����������ʽ�ͽṹ���������¿ɷ�Ϊ6����̬��Ⱥ�������ݡ����ݡ����ݡ����ݡ�½�ݺ����ݡ��־ͳ��������ԼӸ�����
������һ��?Ŀ��Podicipediformes���еȴ�С�����ݣ�����DZˮ����Ҫ����Ϊ��ֺ�߷���İ�״�롣��̶��ۡ���ë������˿�������ϵ�ë����Ʒ��β��Ϊ���ɡ���ˮ����ֲ�ᆬҶӪ������
������Ŀ���༫��DZˮ������ʱ�ܱ���������ˮ��DZ�ӣ��������������ݲ�ͬ��DZ����ɳѼ�����ɱ���������ˮ�����ӣ����ҹ���������ΪС?��Podiceps ruficollis����ͼ19��30��������ˮ��«�����С�Ƹ룬����Һ�ɫ����Ϣ��ˮ�ݷ�ï�ĺӺ��ڡ���ʳ�ԣ�����ˮ������Ϊ������ֳ������ˮ�ݣ���ɸ�������������ɫ���ɫ����Բ���ѣ� 6��9ö�����۾��μӷ��ѣ�������18�� 29�졣�ڷ����ڼ䣬�����볲ʱ���������ˮ�ݽ��Ѹ��ǵ�ϰ�ԣ���Ѽ��Ҳ�ߴ�ϰ�ԣ����б��¼����������á������ҹ��Ĺ۲��Լ����ⱨ�������ָ���ϰ�Կ����Ƿ������Σ����һ����Ӧ���ҹ�����ѻ���ر���С����ѻ������Ÿ�Լ�����������Σ���Ѻͳ������Ҫ��С�

��
��������������Ŀ��Procellariiformes����������Ŀ�����ͺ��������ࡣ�����ƺ�Ÿ�������ʹ�׳�������峤�ɽ�1m������Ҫ����Ϊ����ǿ��߹����ɶ�������Ƭ�����ǡ��ǿ׳ʹ�״��ֺ����롣�������⣬���ڰ��衣�����ڰ��ߵĵ��ϻ�Ѩ�У���ʱ�����Ե��Բ�Ҷ���Ѱ�ɫ��ÿ��һö�����Ծ��μӷ��ѣ�������70��80�졣�������貸��42�죬Ϊ������
�����ҹ��ϳ������������β�����̣�Diomedea albatrus����������ΪƯ���Ժ�����ֳ���⣬�����հ������Ϣ�ں��ϡ���־���ؿ���8000km��Ǩ�ɣ�����“�������”�ļ�¼��ͼ19—31����
��
��

��������������Ŀ��Pelecaniformes���������ݡ���Ҫ�����У�4ֺ���һ������Ĥ��ȫ�룩����ǿ��߹������߷���ĺ�������Ӧʳ���ϰ�ԡ��ҹ����������а������ɣ�Pelecanus philippensis������ˣ�Phalacrocorax carbo����ͼ19—32����
����������Ҫ�ֲ����ȴ����´�������ͨ�����ɫ�����𰵺ڡ������ر�Դ��ݴ沶���������������ɢ�����¡��dz��ڼ�ȡʳ�س����ڴ�����¶�����ң������Ⱥ����ͷ���ھ�ʳ��ӪȺ�������ϻ�ر�������1��4ö��ɫ�ѡ����Է���Լ6�ܣ����������貸��35�졣
��������ֳ���ӥ��ͨ�����ɫ�����л��ߡ������ɴ�Ⱥ���㣬��������ʱ�ɶ���ҵ���Σ������ȺӪ�������ϻ��ҷ��У���2��4ö����ɫ�ѡ����Է��ѣ�������25�죬�������貸��21�졣�ҹ��Թż�ѱ����˲��㡣�����ڴ��ַ�������ҵ��Դ��Σ������ź��Ѳ����á�
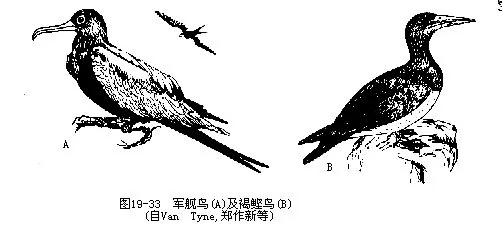
����С������Fregata minor����������Sula leucogaster��Ϊ�ҹ���ɳȺ�����������࣬�����ȡ��´����������ࡣǰ�����Ӷ������࣬����������ݵķ��У��ڸ߿ն�ʳ�������ɡ���˺ͺ�Ÿ�������������ࡣ�������Ļ�������ʱ�����������˲���ڰ��н���ˮ����������ʳ������ϰ�����������Ǻ����ģ�ͼ 19—33����
��
�������ģ�����Ŀ��Ciconiiformes�����������ݡ�����ˮ�ߣ���ˮ����졢�����Ⱦ������ֲ���¶��ֺϸ����4ֺ��ͬһƽ���ϣ��˵�����ͬ�����������ɡ�
��
�����ҹ������������࣬�������ء��������κ����ƣ���ǰ����ֺצ�ڲ����״ͻ����������������“ S”�ͣ�ͼ 19—34����
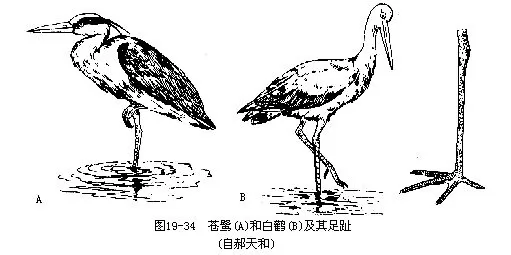
�����ҹ����������к��٣�Ciconia nigra�����������٣�Ciconia boyciana�����ڸ��������·�϶��Ӫ������3��5ö���ѣ�������30��38�졣�����������ҹ�������ֳ��Ϊ�������������ݣ�Ӧ�ϼӱ�����
�������ೣ���ּ�ȺӪ���������������泤��һ�������“��”���ܲ��ϵ�����������ɷ���״��������ʳ��ʱ��ճ�ŵ������ֺ����״�ἴ��������������ҹ����������д���أ�Fgretta alba�������أ�Ardea cinerea�����������ҹ����ж��֣�����������࣬�䴿��ì״�������Ϊ���ص�װ��Ʒ��������Ҿ�����ȡ����ʹ���������½���Ӧ������������ȥ���ú��ڵĸ����ϼ��а���Ӫ�����������ª����3��5ö����ɫ�ѡ�
�������壩����Ŀ��Anseriformes�����������ݣ�����Ҫ�������ࡣ����Ҫ����Ϊ����⡢��Ե������״�ΰ壨����ʳ���ܣ�����˾Ӻ��“���”���Ⱥ��ƣ�ǰ3ֺ����롣���ķ����ϳ��з��������ɫ����ɫ���ɫ�İ߿飬��Ϊ“����”�����ܻ��������Ĺ����ң������ڷ���ʱ�Ĺ���������߽������١�β֬�ٷ������������貸�����ܶ�����������
��������Ŀ����鲼��ȫ���磬��Ҫ�ڱ�����ֳ������м����Եij�����Ǩ���ϰ�ԣ��������ҹ���ֳ����·��Խ����������40���֡�ͨ����˵��ҰѼ���㼰��������Ŀ��ͼ19—35�������������У�
������ͷѼ��Anas platyrhynchos��������Ѽ��Anas poecilorhyncha��Ϊ������Ѽ�࣬�����ҹ���ֳ����Ѽ���Ǵ�����ѱ�������ġ���ͷѼ������ɫ������ͷ�;�����ɫ�����²��а����������Һ�ɫ��������غ�ɫ������Ѽ���۾����غ�ɫ����ں�ɫ���ȶ˾ߵ���ɫ�߿顣��ֳ���ҹ��ӱ�ʡ�����Ա�������Ӫ���ڰ��߲ݴ��У����Ӳݼ������ģ������̻�ɫ��8��12ö��
���������������к��㣨Anser cygnoides�������㣨Anser fabalis�������ͽ�Ѽ�������ɫ���ƣ�����ɫΪ�����������ͷ�����������ͷ�̡�����Ϊ�ҹ��Ҷ��ԭ�档�����һ������ϰ����Ѽ����½���Խ�ǿ����ֲ��Ϊ��ʳ����Ǩ�㼰Խ���ڼ䣬��ʱ�Զ�С��Σ�����أ������ü�ֵ��Ϊ��Ҫ���Զ���
��������Ŀ��������������죨Cygnus cygnus����ȫ���ף���ƾߺڰߣ���Ӿʱ����ֱ����ˮ�档����������������Ϊ������Ʒ�����⡣����ϡ�ٶ����Ϊ�ҹ��ص㱣������֮һ��
��������������Ŀ��Falconiformes����ʳ�����࣬�������͡����������˺�Ѳ������ǿ���������������Ĺ�צ˺ʳ���ࡢС�ޡ��ܡ����������ȶ���Ƽ��ɼ����裬�����������������ԡ�������������������
�����ҹ�����Ŀ�������༰�������࣬��ë���ر��Ƿ���β������Ҫ���ü�ֵ�����������к������Falco vespertinus�������ʹ�С�Ƹ룬�������ɫ���Ƚź�ɫ�����п�����࣬���������ӡ����ļ������ҹ���ֳ����ռϲȵ����ѻ�����ѣ�“ȵ���ռ”��“�”��ָ���������Ӫ��ϰ���ϵ��ص����ټ��ģ�����ʳ�����棬������������𰣨Milvus migrans��Ҳ�dz����Ͽ�ϰ�������ݣ������ɫ��β���״���ʣ�Pandion haliaetus����һ��ר������Ϊʳ�����ݣ�����ֺ�ܺ�ת����ֺ�¼�צ����߷�������״ͻ��������ץ�����ࡣͺ�գ�Aegypius monachus��Ϊ�ҹ����ڵĴ������ݣ���Ҫ��Ϣ���ҹ������������ĸ�ɽ�ϣ���ʳ����ʬ�壬ͷ����ͺ���������Ϊ���ද�����Ҫ������ͼ19—36����
��������Ŀ�������ë���о��ü�ֵ�⣬��ʳ�Զ��ۣ��������������ಶʳҰѼ�����ݺͼ�����⣬�������������Ϊ��ʳ��������ũ��������������������Σ�����棬������̬ѧ����Ϊ������Щ���ݵ��������ܶȣ������ģ���������ھֲ�������ʱ���ڹ�����в����������ץ��IJ����������������У��������ڶ������Ⱥ����������ȴ���л������á��������Ŀ������������ѱ������ҹ��ص㱣������
��������Ŀ�쳲�ڸ��������·�϶�ڣ���1��6ö�������ε��ѡ��Ѱ�ɫ����ɫ����������ϡ�費�ȵĺְߡ������ɴ����λ������Ը�����������Լ35�졣�׳���鲼��ɫ��ë�����貸��40����ʼ�볲��
�������ݾ����³�“ʳ��”��ϰ�ԣ���������Ϣ�أ��ر��dz��Ľ��ԣ���Ϣʱ��������������ࡢ����ȵIJ��������IJ��ţ��ر��ǹ�������ë���³����ɼ��ͷ�����Щʳ�裬���ڲ��������к������������������䶯�����ṩ���м�ֵ�����ϡ�
�������ߣ�����Ŀ��Galliformes����Ӧ��½�ܲ��У�����Ŀһ�𱻹���½���ࡣ�ȽŽ�׳�������ھ�����ʳ�Ķ�צ�����칭�Ρ�������ʳֲ�����ӡ���ҷ������Բ������Զ�ɡ����۴����ɫ��������ɫ���ޣ���ֳ�ڼ�ö������и��ӵ���ż��ҫ��������ɡ�
��������ĿΪ��Ҫ�ľ������࣬���⡢�����⣬���кܶ�����Ϊ�����Ĺ����������в������ҹ��ز����ҹ�����Ŀ����ʮ�ַḻ�����Ҵ��������Ϊ�ܶ���Ҽ�������������������������Ժ��ϸ�Ļ�������ʩ�Ǽ�Ϊ��Ҫ�ġ����ҹ���������������Lagopus lagopus����黼���Tetrastesbonasia���DZ������͵Ĵ��������ǵ����Ų�������ë���ࣨ�Ⱥ�Ĵ�״����Լ��ǿױ��𣬿�����������������������Ӧ��̦ԭ�ش������ֳ����ë��ɫ��������Ϊ��ɫ����ѩԭ����������ౣ��ɫ�������������̿�ȸ��Pavo muticus�������£�Lophura nycthemera�����츹����������Chroysolophus pictus������������Phasianus colchicus������β����Syrmaticus reevesii����ԭ����Gallusgallus���ȣ���Ϊ�о��ü�ֵ���ɹ����͵����ࡣ��������Crossoptilon mantchuricum��Ϊ�ҹ��ز���ϡ�����࣬���ںӱ�������ɽ�������ľֲ�ɽ���У�����Ҫ�ı�������ԭ���ǼҼ������ȣ��ҹ�����ѧ����֤���Ҽ����ҹ��Թž���ѱ������ν“�й��Ҽ��Ǵ�ӡ������”��˵��������ȷ��
��������Ŀ������һЩС�Ͷ������࣬�����������ȣ�Coturnix coturnix������Francolinuspintadeanus����ʯ����Alectoris chukar���ȣ����ǵ����Ը�ǿ����Ϊ�������ߵ�������ͼ 19��37����
��������Ŀ��ֲ������Ϊ��ʳ������黼�����������֦ҶѿΪ��ʳ������ֳ�ڳ�Ϊһ�۶�ƣ���������ڵ���Ӫ����һ���8��15ö�ѣ���ɫ������������20��30�졣
�������ˣ�����Ŀ��Gruiformes�����ʹ�С���ȵ����ݡ���������ȡ������ϳ����ֲ�ͨ����¶����ֺ����������룬4ֺ����һƽ���ϣ���ֺ����ǰ3ֺ��������Ŀ����Ϊ�����
������Ŀ���������������ף�Grus japonensis��������ϡ������֮һ�����ҹ����������ɹ�������������ֳ�����ҹ��ص㱣�����ࡣ�������߿Ժ����������ķ������йأ����������¶��̾����عǸ�������������������죬�Ϻ��̳ɶ�Ȧ���������ع��ڡ�Ӫ���ڵ��棬��2�ѣ����Է���32�졣�������Ƭ�̺��볲����ʱ����Ϊ����İٷ�֮һ���������������ǽ�Ϊ�ټ��ġ�
������������Ƕ���Fulica atra��Ҳ������Ŀ���࣬���������ڶ࣬Ϊ���Զ���𱣨Otis tar-da�����׳Ƶ��L�����ܷ�����������������ģ����ڲ�ԭ�ĵأ��Ա���Ϊ�������ҹ����������ɹ���������ֳ���������ڹ����Բ����Ѻ�ϡ�٣����磬Ӣ����1838�꼴�Ѿ��������������Σ���֡��ҹ��ص㱣�����ࣨͼ19—38����
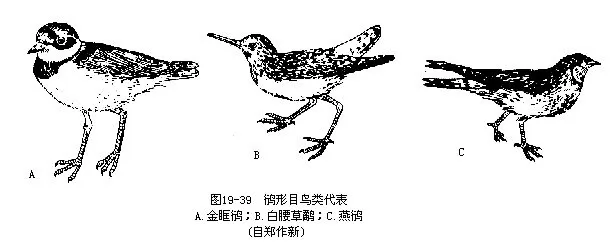
�������ţ��a��Ŀ��Charadriiformes�����ݣ���Ϊ��С�����࣬����ܶ࣬��Ҫ�ֲ��ڱ��������Ϊɳ��ɫ�����ܿ��١������Ʒɡ�����Ϊ�����������ɫ���������ԡ���ͻȻ��ɶ����������˶����Ե���Ҫ�����еķ���ϵͳ��Ÿ��Ŀ���뱾Ŀ��
�����ҹ����������н���a��Charadrius dubius����ΪС���a�࣬�ں�̲�ϲ���������4ö����ɫ��ɰʯ�����ƣ������֮��Ҳ�ѷ��֡�������Լ20�졣�������֣�Tringa ochropus��Ϊ����ij���������ͨ����˵“�ְ����������˵���”������ָ���������a��Glareola maldivarum����̶�����β�ֲ棬���������ӣ�Ϊ�ҹ������IJ�ʳ�ȳ�����ࣻ����ɽ��ɽ���������飬һ�����aһ�����ڿ�����ȳ���16200����ͼ 19—39����
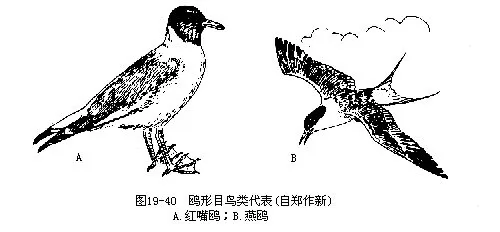
������ʮ��Ÿ��Ŀ��Lariformes�����������࣬���a��Ŀ��Ե��ϵ���У���ϰ�Խ������ݡ�����Ϣ��ˮ�߲�ʳ���������ݡ�������Ϊ����ɫ��ǰ3ֺ����롣���ⳤ���Ʒ��衣��������̬��Ϊ���������������������ϰ�������������ڵر�����2��4ö�������ѣ�������Լ20�졣
�����ҹ����������к���Ÿ��Larus ridibundus������Ÿ��Sterna hirundo�����������Ƹ�������࣬�����뵽��½��ֳ��ͼ 19— 40����Ÿ����Ÿ�����ɴ�Ⱥ�������ҵ����ʱ�ɶ��������Σ��������Ÿ���׳ƺ��ࣩ�Ķ���������ʳ�ݵ����Ⱥ��棬Ϊ����ҵ��������
��
��
������ʮһ������Ŀ��Columbiformes��½�ݡ���̣���������������ǿ��������Ĥ��cere������ȴ��׳��4ֺλ��һ��ƽ���ϡ���ҷ�����������ܷ��ڸ���ι��������Ϊ������Ŀ������Ϊ���Զ���

 ��
��
�����ҹ�����������ë��ɳ����Syrrhaptes paradoxus�������������ڼ����֮�䡣���ڻ�Įɳ�أ������ɹ���������ֳ����ɳ��ɫ����ֺ�˻���ǰ3ֺ���ϣ�����ë�������Ʒɣ�������ǧ��ֻ��Ⱥ�������ɵ�Զ����Ǩ�ɣ�Ѱ��ˮԴ��1888���ҹ�ɳ����Ǩ������Ӣ���������ڵ��棬�Ѱ���ɫ�ߺְߣ�������Բ�Ρ�������28�졣����Ϊ��Ҫ���Զ���ԭ�루Columba livia��Ϊ�Ҹ�����ȣ��ֲ�����ȫ���ҹ������Ͻ������зֲ���ɽ��楂Streptopelia orientalis�����龱��楂S.chinensis����Ϊ���������ࡣӪ�������ϻ���ʯ�죬��2ö��ɫ����Բ���ѡ����Է��ѣ�������14��19�졣����������12��18�죬��һ�����������Ը��飨ͼ19��41����
 ��
��
������ʮ��������Ŀ��Psittaciformes�����ݡ���4ֺ��ת���ƶ�ֺ�ͣ������Ӳ����������������̧������������������Ե��������Ƥ�����Ӫ���������С�����ɫ�������ѣ�����21�졣����Ϊ������
��������ĿΪ�ȴ����࣬�������Ĺ������ҹ����ϡ������ļ��ϲ��Լ����ϵ�������������ģ�Psittacula alexandri��������Ŀ���������⣬ԭ�����Ļ�Ƥ���ģ�Melopsittaccs undulatus���ѹ㷺��Ϊ������������Ŀ�����ж����Թ���Ϊʳ��������ʱΣ�����ء���Ч���������ĵ�����ϰ�ԡ�
������ʮ��������Ŀ��Cuculiformes�����ݡ���ֺ�͡���������Сӥ�����졢צ���߹��������ֲ���ŷ�Ǵ�½������Ϊ�����Է�ֳ�����Ѳ��������С�����Ϊ������
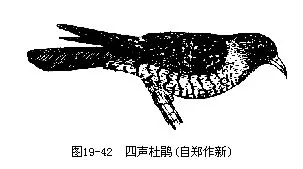
�����ҹ����������д�ž飨Cuculus canorus���������ž飨C.micropterus����ͼ 19��42����ǰ�߽�����“����”�����ֳƲ������߽�����“������”�����ڶž�Ĺ�����Ǩ��ʮ��������ÿ���紺������������������ҹ��ͣ������Թż����������ǵ�ע�⣬����Ϊ����ʫ�衢��˵�����⡣
������ž����ȸ��Ŀ���ࣨݺ�ơ�ȸ�ơ���ü�ơ��ơ����Ϳơ�ɽȸ�ơ�ѻ�Ƶȣ�Լ100��������ij��в��ѣ�����������ɫ�������Ѻ������ҹ�����Ĵ�ž������Ǵ�έݺ���׳������磨��Լ13�죩�����Ǻ��������ı��ܣ��ܽ����������Ѻͳ��׳����⣨ͼ19—43�������������IJ������ž�ĿΪ����������ʳ��ë�棬��������ʳ���������Ȳ��˵ġ�
��
������ʮ�ģ��^��Ŀ��Strigiformes��ҹ�������ݣ��ڲ��ṹ�������ݡ������ξ߱��������������⣬����ֺ�ܺ�ת�ɶ�ֺ�ͣ�������Ե�����۴����ǰ�������з���״ϸ�ɵ�“����”��������ҹ��ֱ����졣����Ϊҹ�����Ҫ��λ���٣������ش�����Ե�����Ż�߶����������ռ���������Ƭ��������ʱ������Ӫ���������У�����ɫ������1��7ö������Ϊ������
�����ҹ����������糤���^��Asio otus����ͼ 19—44����������è������“èͷӥ”���^�ೣ����ҹ���������������Ľ�����������ò�����������ۣ���˳�����Ϊ��һ�ֲ���֮����ʵ����������ʳ������ʳ���е�90������Ϊ���ࡣ������ҹ������ı��죬Ϊ����Ŀ�������������^�������Ϊ�ҹ��ص㱣�����ࡣ
������ʮ�壩ҹӥĿ��Caprimulgiformes��ҹ�������ݡ�ǰֺ�������ϣ��Ʋ�ֺ�ͣ�����צ����״Ե����Ƭ��������ʱ�������ڿ�������Ե�߳���Ӳë����Ӧ�ڷɲ����档��ɫ���֦ɫ���ƣ�Ϊ����DZ��ʱ�ı���ɫ��ͼ19—45������Ӫ������ 1�� 2ö���ڵر�������Ϊ������
����ҹӥ��ʳ�ó棨����һ��θ�м��500��ֻ�ó棩����������ĸ�������ž�֤ʵ��ҹӥĿ����������������Զȹ�ȱʳ�ĺ��伾�ڡ��ҹ���������Ϊҹӥ��Caprimulgus indicus����
����ҹӥ������ѧ��Ʒ����˵��“ҹݺ”������ΪС�����ݣ������ҹ������������������Luscinia calliope�����Ƶ����ࡣ
������ʮ��������Ŀ��Apodiformes��С�����ݣ�������������Ƶĺ�ֺ��ǰ����ǰֺ�ͣ������߹�����Ϊ������

��
�����ҹ���������Ϊ¥�ࣨ�������ࣩ��Apus apus���������ɴ�Ⱥ�ڸ߿ռ��ɲ��棬�������Ƽ��࣬��4ֺ��ǰ����˿�ࣨCollocalia spp.����ֳ������Һ�ٷ�����Ӫ�������������̲�Ʒ“����”��
��������Ϊ��������С�����࣬���ؽ�1g���ң���Ҫ�ֲ��������ޡ��Ի���Ϊʳ�����ڻ�ǰ��ֱ���ɻ����“��ͣ”����ʱÿ����������50��Σ����ؼ���Դ�СΪ����֮�ڣ�ͼ19—46�������귢�ִ����ڻ��۶�ȱʱ�������ߵ�������
��
������ʮ�ߣ���ɮĿ��Coraciiformes�����ݡ���Ϊ��ֺ�͡�����϶࣬������졣Ӫ��������Ϊ��ɫ�����ѣ�����Ϊ������
������Ŀ���ҹ��ij��������д���Alcedoatthis���ʹ�ʤ��Upupa epops�����������δִ����䣬�������ɫ��β���С������ϺΪ��ʳ���ذ��´�����ѨΪ������ʤ��ϸ�������䣬�Ե�������С����Ϊʳ��ͷ������״�����׳ƻ����ȡ����������������϶�Լ���ѷ����Բݾ�����״������4��8ö��Բ���۰�ɫ�ѡ�������12��15�졣�����ڷ����ڼ䣬����β֬�ٷ���һ���س��ĺ���ɫҺ�壬�Գ������б������á�
����˫��Ϭ��Buceros bicornis��Ϊ�����ҹ������ϲ������������ࡣ�������䣬���춥���н���¡����ڸߴ������������������ڴ�����ڶ��ڣ��������������������Դ�θ��Ż���ķ����ォ���ڷ�գ������Կ�������Ķ�϶���Խ�������ιʳ������������У�������ߵȣ��˺���һ����Ӧ�������� 28�� 40�졣ֱ����������ʱ������ʼ“���Ŷ���”��ͼ 19��47����

������ʮ�ˣ�?��Ŀ����ľ��Ŀ����Piciformes�����ݡ���Ϊ��ֺ�͡��������䣬רʳ��Ƥ���ܾӵĺ��棨����ţ�׳棩��β���β���Ӳ�����е��ԣ�����ľʱ����֧�ܵ����á��䶴Ϊ������3��5ö��ɫ����Բ���ѡ�������10��18�죬����Ϊ������
��
������ľ��Ϊ������ɭ�������������е�������Ƥ����ĺ���֮�⣬���ɸ�������ľ�ĺۼ���Ϊɭ��“�����ɷ�”��ָʾ�����������Ϊ“ɭ��ҽ��”���ҹ����������а���ľ��Picoidesmajor����ͼ 19—48����

��
������ʮ�ţ�ȸ��Ŀ��Passeriformes�����ݣ�ռ�ִ�����ľ��������Լ5ǧ���֣�������Ҫ����Ϊ�����ܼ��������ӣ�������������ֺ3ǰһ��ֺ����ֺ�ȳ�������ֺ�ͣ������ź���Ƭ���ϳ�һ���������۰塣���Ӫ���������Ϊ������
����ȸ��ĿΪ��������ߵȵ���Ⱥ���������������ʷ�Ͻ�������Ŀ�������������ھ��ҵķ�������Σ�����ࣨ���64���ƣ������������а��飨Melanocorypha mongolica�������ࣨHirundo rustica����ϲȵ��Pica pica����ͺ����ѻ��Corvus frugilegus���� ��ü��Garrulax conorus������ü��ݺ��Phylloscopus inornatus������ɽȸ��Parus major������ȸ��Passer montanus������ȸ��Fringilla montifringilla���ȡ������е���ũ�������е���ũҵ�����е���������������Ч����Ϊ����������ͼ19—49����
��

������ ����ķ�ֳ����̬��Ǩ��
��
һ������ķ�ֳ
��
�������ֳ�������Եļ����ԣ����и��ӵ���Ϊ������ռ�������������ѡ������ȣ�����Щ���������ں��������Ӧ��
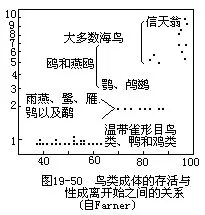
����������Գ�����������һ�꣬�������ݼ�Ѽ��ͨ������һ��ʹﵽ�Գ��죬�����ȴ�����ʳ����������3��5���¼��ɷ�ֳ��Ÿ���Գ�����3�����ϣ�ӥ��4��5�꣬�����̼�أӥ����9��12���Գ��졣�Գ��������һ����������Ⱥ������������أ�ͼ19—50�������������͵ģ��Գ���������ÿ������ֳ�ij�����Ҳ�١�
��
����������������ż��ϵά�ֵ���ֳ�����ˡ������볲Ϊֹ����������Ϊ������ż����֪������졢��졢�㡢�١��ס�ӥ���^�����ġ���ѻ��ϲȵ��ɽȸ�ȡ������������У���2���ƺ�4���ǿ�������һ�۶�ƣ������ɼ���������������֯����Լ0.4���Ƽ�1���ǿ�������һ�ƶ��ۣ�������ֺ�ȼ����֣���������Ϊһ��һ�ơ�
������ͨ����ÿ�귱ֳһ�ѣ�brood������������ȸ��������ȣ�һ��ɷ�ֳ���ѡ���ʳ��ḻ���������˵���ݣ����ֳ��������ÿ�ѵ������������ࡣһЩ�ȴ�������ʳ�����������������귱ֳ��
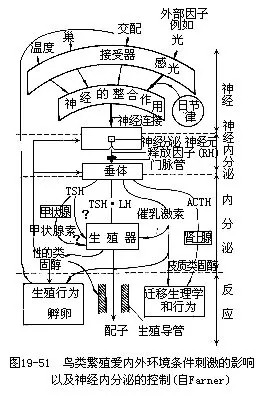
�����������ٵķ����ͷ�ֳ��Ϊ�ij��֣�����������������£�ͨ�����ڷ���ϵͳ�ĵ��ڼ���ʵ�ֵġ�ÿ�괺�������������ĸı��Լ��������۵ı仯�����ӣ�ͨ������ĸй���������ϵͳ��Ӱ�������²���˯�����࣬ʹ���ദ���˷�״̬�������²���������Ԫ��������Ԫ�������´����ž����ڷ����ͷ����ӣ�RF�����������´�����ڡ����´��������ڵ����ݴ̼��أ�FSH���ͻ��������أ�LH����ʹ�ѳ�����ϸ�������������Լ��أ��Ե���̴�����ʹ��ֳϸ�����첢����һϵ�з�ֳ��Ϊ�����´��������ڵĴټ�״�ټ��أ�TSH����ʹ��״�ٷ��ڼ�״���أ��������л���Ĵ�л��������ֳ��Ϊ�������ԡ����´��������ڵĴ�������Ƥ�ʼ��أ�ACTH����ʹ�����ٷ����������أ�������л�������̼���Ӧ������������������뷱ֳ�йص�Ǩ�����Ϊ��ͼ19—51����������������ֳ�����ڣ��������ż��ҫ�����䡢�쳲�ͷ��ѵ�һϵ�л��Ҳ�����ϵ�ͨ���й����������ڷ���ϵͳ��ǿ�������������ڵ����������Ϊ���ս��ɣ���ҹ���ɣ���circadian rhythm�������������ӣ��Է�ֳ���ڻҲ��Ӱ�졣
��������ÿ����뷱ֳ�����Ժ��������ٵķ���������һϵ�еķ�ֳ��Ϊ��������ֳ����Ǩ�㡢ռ������ż��ҫ�����������Ѻͷ����Լ�������ȣ��������볲֮������ʼ�^����½���뿪Ӫ���ص㣬�����˵ĵ���Խ�����־�һЩ��Ҫ���ݼ��Խ��ܣ�
������һ��ռ��������territory�������ڷ�ֳ�ڳ�����ռ��һ�����������������ࣨ������ͬ�����ࣩ���룬��Ϊռ��������ռ�е�һ����س�Ϊ����ռ������ż��ҫ��courtship dis-play������ԣ�pair formation�����л��ؽ����һ��ģ�ռ���ɹ�������Ҳ����ż��ҫ��ʤ���ߡ�
��
����ռ��������ѧ������Ҫ�����ڣ��ٱ�֤Ӫ���������ھೲַ����ķ�Χ�ڣ���ó�ֵ�ʳ�﹩Ӧ�����Է������������ġ�ʳ����Դ�����ḻ���ȶ��ģ��Լ������漰����Ϊʳ�����࣬������ı������������ڵ���Ӫ��������������Ⱥ���ܶȺͷֲ���������Ч��������Ȼ��Դ���ֲ��������ܼ���Ҳ�ɼ��ٴ�Ⱦ����ɢ�����ۼ��������������ԡ������������Լ����ѡ������Ȼ�ĸ��ţ��ܶԸ����μӷ�ֳ��ͬ���������������Ӱ�죬��������Ե��˷����á�
��������Ĵ�С�ɴӼ�ƽ��km2������ӥ���m���ա���ѩ�^��������m3��������һЩȸ��ĿС�������ԼΪ����m2�������С�ǿɱ�ģ���Ӫ�������˵������ޡ���Ⱥ�ܶ���Խϸߵ�����£�����ɱ���������“ѹ��”��“�ָ�”����С�������ҹ�����������������ǧ��Ŀ���������ɭ�ֱ��ı�Ϊ���أ������Ѽ�����С����ijЩȸ��Ŀ��������Ϊ���ԣ��������ǿ����ƶ�����ռ���쳲���ݻ�;���������ǣ����ڻ��������ĸı䣬���˳�ַ���ޣ�����ʹӪ“����”�����౻��ѹ����������“��ɢ��Ⱥ��”���ٽ�һ��ѹ�������γ�“Ⱥ��”��
����������ռ����Ӫ�������У������Բ�ͬ�̶ȺͲ�ͬ��ʽ����ż��ҫ��ͼ19—52�������������������У�����ȸ��Ŀ��Ϊͻ��������ż��ҫ�����ж���ʹ��ֳ�����˳�����еı��ܻ��ʹ��ϵͳ���ڷ����ٴ��ڻ���״̬���������Ե��Ի���Ӷ�ʹ���Ե������ٷ���������Ϊ�ķ�չ����ͬ����synchronize������ż��ҫ�������Եı��ϣ��ر��Ǵ���ͬ�����ࣩҲ��ʮ����Ҫ�ġ�������Ϊ��������ż��ҫ���������У��������д������ֵ������ԣ����������Ե��ϵ�Ͻ��IJ�ͬ�����࣬��������ѧ�ĸ���������ã��ɱ����ּ��ӽ�����ż��ҫ�˥�ˣ����������µ�“������”������ʱ�������·�ֳ�����жϡ�
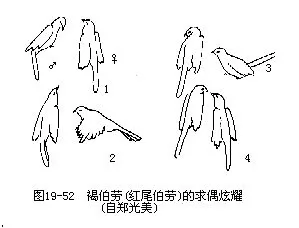
��
����������������nest-building����������������������Ϊ���͵�������ڵر���Ѩ�ڷ��������ݡ���Ҷ��ë���ߵ����ࣨȸ��Ŀ������ϸ֦���ݾ���ë����ȱ�ɸ�ʽ�������µ����������¹��ܣ���ʹ�Ѳ��¹�ɢ����ͬʱ���������������ڱ��£���ʹ���ѳ����Ѽ�������������˺�������Ӫ���ɷ�Ϊ“����”��“Ⱥ��”���ࡣ����������Ϊ���������ɢ��Ⱥ����Ⱥ���ڵ��켰�˼������ĵ�����Ϊ��������������ֺ�����졢�����̡����ɣ���Ÿ�ࡢ���ࡢ�����༰ijЩѻ�����ࡣ���༯ȺӪ���������ǣ�������Ӫ���ĵص����ޣ���Ӫ��������ʳ��ȽϷḻ������������׳�����Ҫ���������ڹ�ͬ������С���Щ�����У�����“����Ӫ���ص㲻��”����Ҫԭ�������������Ȼ��Ĵ��ģ���������˳�ַ�Ľ�һ�����٣���ȺӪ�������ƽ��������ԡ�
�����ҹ�������������ṹ�ص㣬�ɷ�Ϊ���¼��ࣨͼ19—53����
����1.���泲��ȥijЩȸ��Ŀ���ࣨ����顢��ݺ��Ҳ���ڵر���֯���ɵij����⣬���泲�����ŵ͵ȵ��ܻ�ˮ�����ࣨ��������Լ���½�ݡ����ݡ����ݣ��ij�ʽ�����Ľṹ��ª����ɫ�뻷�������ƣ���������Ҳ��ͬ���ı���ɫ��
����2.ˮ�渡��ijЩ���ݼ������ܽ�ˮ�����۲���ɺ���״������ˮ��������
����3.����������������������϶�ڣ�һЩ���ݡ����ݼ�����ȸ��Ŀ�������ˡ���Ѩ��λ�á��ṹ�����������ϰ�������й�ϵ�����нϵ͵ȵ�������ٸ��ӳ��ģ�����ɫ�ѣ���ӳ��ԭʼɭ����������������ѣ���ȸ��Ŀ�����������ڶ������Ƹ��ӵij��ģ���ɫҲ��������ӳ����һ�ֺ������ػ�����
��
��
��
����4.��֯������֦���ݾ���ë����ȱ�֯�ij����͵����ࣨ�����ԡ����ࡢ���ݣ��ij��ͼ�ª��ȸ��Ŀ�������ܱ�ɸ�����ʽ����״����״��ƿ״���ľ������ҹ����쳲������������֯����Ploceus philippinus���ͷ�Ҷݺ��Orthotomus sutorius����ǰ�����콫ֲ����ά��֯����ش����֯��ƿ״����������ֲ����ά�ᴩ������Ҷ�IJ�Ե������ϳ��������ҵĶ�״����
������������ij��֣��в������ࣨ�ر��Ƕ������ࣩ��ת���ڽ�������Ӫ����������Щ������Ӵ������е����࣬Ҫע���о����溦��
�������������ѣ�egg-laging������ѣ�Incubation���Ѳ��ڳ��ڲ����Է������ѵ���״����ɫ����Ŀ���Լ��ѿǵ����ṹ������Ӿ��������ͬһ��Ⱥ�䳣�������Ƶģ��Ӷ�Ҳ���Է�ӳ����ͬ��Ⱥ֮�����Ե��ϵ������Ϊ�о�����ѧ�����ݡ�
����ÿ�������ڳ�����������������Ŀ��Ϊ��������cluth������������ͬ���������ȶ��ģ�һ��˵�������Ѻͳ��ı��������ơ��ɻ������ߵģ����������١���ͬһ������ԣ��ȴ��ı��´��IJ����٣�ʳ���ʢ��ݵIJ������ࡣ��������������Ȼѡ��������ġ�������������ȵĺ����Ŀ�����⣬������Ҳ����������ķ��Ѱ������ڸǵ���Ŀ�йء�
���������д����Ŷ������ѣ�determinate layer���벻�������ѣ�indeterminate layer���������ͣ�ǰ����ÿһ��ֳ������ֻ���̶���Ŀ����������������ʧ������������롢��Ÿ����������ϲȵ�ͼ���ȡ������������ߣ���δ�ﵽ�������ѵ���������ǰ����������ʧ������һö�����ѻʼ�մ����˷�״̬��ֱ����������е�������Ϊֹ����֪һЩ��졢����Ѽ�ࡢ���ࡢһЩ��ľ���Լ�һЩȸ��Ŀ���ࣨ�������ȸ�����д����ԣ��������ü��ݣ��������ȡ�Ѽ���켰�ȣ�����������������������ԡ�
�������Ѵ��Ϊ�����Σ����粮�͡�Ѽ������ȣ���Ҳ�е�Ϊ�����������ѣ���ھ�β���롢���ٵȣ�����������Ϊ������ѣ�����š���ֺ�ȵȣ��������η����ߣ�����ɫ���ֻ��ƴ����������ࣨ������졢��ˡ�Ѽ���죩֮�⣬������ѵ����������з��Ѱߡ����Ѱ��е����ģ�����ܶ�ȸ��Ŀ���ࡢ���ݡ��뼰?����������λ�ģ����纣ȸ���a��Ŀ���ࣩ�Լ�һ�������������λ�ģ�����Ÿ�뼦�ࣩ�����ѰߵĴ�С������������֮��û��ʲô��ء�
�����Ѳ��������ʱ������Ϊ34.4�桫35.4�����ҡ��ڷ������ڣ������¶ȸ��������¶ȣ�����̥�������ڣ������¶��Ը��������¶ȡ�
����ÿ������ķ�����ͨ�����ȶ��ģ�һ���������ķ����ڽϳ�����ӥ��29��55�졢������63��81�졢�Ҹ�18�졢�Ҽ�21�졢��Ѽ28�졢��31�죩��С����������ڶ̣�����һ��ȸ��ĿС��Ϊ10��15�죩��
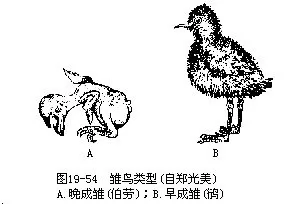
��
�������ģ�������Parental care����̥��ɷ���������ⲿ��ʱ�����Ľ���ͻ��——“�ѳ�”�������ƶ���������ij����Ϊ��ɳ���precocial�������ɳ���al-tricial������ɳ��ڷ���ʱ���ѳ�ַ��������������������ſ����Ƚ�������������ɺ�����������ʳ�����������������������ˡ����ɳ�����ʱ��δ��ַ���������������ϡ�������۲������������������γ���ι���Ӱ���µ�8���²��ȣ��������ڳ�����ɺ��ڷ����������������ͼ19—54����ȸ��Ŀ�����ݡ������Լ�һ�������ݣ�����������͵����࣬�����ɡ������̣����ˡ����Կ�������������Ի������ԣ��dz�����Ȼѡ��IJ�����������ζ���ȫ������������������������࣬������Ϊ���ɳ�����ɳ��ǵ���������߳ɻ��ʵ�һ����Ӧ�ԡ�������ˣ���ɳ���������������ʶ���������ߵö࣬���������ĿҲ�ࡣ
�������ɳ��ķ�����һ�����Ϊ“S”���������ߣ��������ڵ������γɺͿ��������ڹ��ɵ����ʻ��ۺ����������ڣ������ڵ��������Ĵ��ڻ��������ڡ��ڳ��������ڣ���ȱ����Ч�����µ��ڻ��ƣ��迿���������ά�ֳ�������¡����ų����ڲ����ٵķ��������Ⱥ����ڻ��Ƶ������Լ����£����¸��Dz㣩�ij��֣���ת��Ϊ���¡����ҹ���������������·����ⶨ�Ľ�������������µ��������ѡ��γ���Ƭ�ĵ��գ����Ǻ��³��ֵ�ת�ۡ�����ֲ��ͣ�Lanius cristatus lu��cionensis��������������Ϊ������10���䣬�������µIJⶨ���뵼���¶ȼƲ�йֳǻ�¶ȣ����Ϊ��

��
�����ܶ������ɳ���������졢���ɡ������̡����ࡢ���ġ�����ʳ����ȣ����볲ǰ�����س�������Ϊ֬���������¡�������Ӧ�����ڳ���ɹ������������������ɵ�ʳ���ȱ����Ϊ�볲ǰ�ķ��𡢼���ȵ������ṩ�ϳ�ֵ����������ڳ������볲ǰ�Ƶ�����������ľ��������������½�������������ɳ������������һ����Ҫ���ء�
�������ɳ������������ڵ�ʳ���ܴ���������Ϊ��ʳ�����ڴ����������������ũ�֡�
��
���������Ǩ��
��
����Ǩ�㣨migration��������������ר�еı��ܻ��ijЩ������綫�Ƿɻȣ���ijЩ���ࡢ�����ࣨ�纣�꣩�Ͳ����ࣨ�����𡢾���������¹�ࣩҲ�м����Եij��������ס�����������к����뾨��Ǩ�����ɴ�����km����ǧkm��������Ϊ����������Ⱥ��˵�������Ǩ�������ձ��������עĿ�ģ����������һֱ��Ϊ����ѧ�о���һ����Ҫ����
���������Ǩ���ǶԸı��ŵĻ���������һ�ֻ�������Ӧ���ܣ���ÿ���ڷ�ֳ����Խ����֮��������Ե�Ǩ�ӡ�����Ǩ�ɵ��ص��Ƕ��ڡ�������Ҷ༯�ɴ�Ⱥ�������Ǩ��������ϱ�����֮�䣬�����ڶ�������֮�䡣
������������Ǩ�����ص㣬�ɰ������Ϊ����resident���ͺ���migrant��transient�����������������ڳ����أ���ֳ������������Ǩ�㣬������ȸ��ϲȵ�ȡ��������ڴ��������������Ź̶���·�ߣ������ڷ�ֳ����Խ������֮�䣬�ҹ������ĺܶ���������ں��������ļ�������ֳ��������ȥ��������ĺ���summer resident��������ࡢ�ž飻��������Խ����������ȥ��ֳ������ƶ�����winter resident������ijЩҰѼ�����㡣�ļ����ҹ�ij���Ա���ֳ���������ҹ�ij������Խ�������ڴ��^�ڹ����Եش��ҹ�ij��·���������������·���缫����ݺ�ȣ���
�����ϸ��˵���ֽ���˵�������в��������ﶬ���ھ���Ư�����ε������ʣ��Ի�����˵�ʳ�﹩Ӧ�����˳�������ΪƯ��wanderer����
������һ��Ǩ���ԭ�� ��������Ǩ���ԭ��ܸ��ӣ��������϶��Ľ��ۡ����������ѧ����Ϊ��Ǩ����Ҫ�ǶԶ�������ʳ��������һ����Ӧ����Ѱ��Ϸḻ��ʳ�﹩Ӧ������������Ϊʳ��������Ϊ���ԡ����⣬������Ϊ�������ļ��ij����գ��糤ҹ�̣������������Ը����ʱ�䲶����ι��������2����������ศ��ɣ����ǻ����ܽ����й�Ǩ�㷽�����漰�ĸ��ָ�����ʵ��
�������˴ӵ�����ʷ���Ʋ�����Ǩ�����Դ���⡣�����������ļͣ�Լ10����ǰ����������α����˶����Ա�����������Ϯ����������֮���������䣬��ѩ��أ��������������档���������Ե���Ϯ����ȴ��ʹ�����γ��˶����������������Ŵ����ܡ���������ʶ���������2�ֻ�������ļ�˵�����ֽ�ķ�ֳ���Ǻ���Ĺ��磬��������ʱ��ʹ����������ȴ�����Ŵ��ı����Դ�ʹ��Щ�����ڱ����������ط����磬����������϶��γ�Ǩ�㱾�ܣ����ֽ��Խ�����Ǻ���Ĺ��磬���ڴ�����ֳ����ʹ������չ�ֲ���������ȴ���������ȥ�����Ŵ������Դ�ʹ��Щ����ÿ���Է���Խ���������磩��
��������˵�����ܽ�����Щ����Ϊʲô��Ǩ�㡣��������ָ�������ڣ����ļ���������ռ����������ʷ�İٷ�֮һ��������������Ŵ��Ե�Ӱ���վ������ġ�����Ҳ�������ų��ڱ�������ǰ���༴�Ѵ�����Ǩ�����ʵ��
����������Ǩ������� ���в��ټ�˵����������Ǩ����շ����أ�������Ϊ���ա�ʳ������Լ�ֲ����ò�ĸı䣬����������Ǩ�����ֽ�ϴ�����ʵ��֤�������������ĸı䣬����ͨ���Ӿ�����ϵͳ�������ڼ����²���˯�����࣬�������ദ���˷�״̬����̼�����ǿ�����´���Ļ���ٽ����ٷ�����Ӱ���״�ٷ��ڣ���ǿ��������ʴ�л����һ����߶����̼��������ԣ��Ӷ�����Ǩ�㡣
����������Ϊ��Ǩ���Ƕ��������̼�������������Է���������������ʷ���γɵ��Ŵ�����Ǩ���“����”�����̼�Ϊ����Ǩ���“����”��
����������Ǩ��Ķ���Orientation�� Ǩ����������ص���ÿһ���־�����Ϲ̶��ķ�ֳ����Խ����������֮��ľ��������km��ǧ��km���ȡ�����ʵ��֤�����ܶ����ࣨ������ࡢ��죩���괺��ɷ���ԭ����ֳ����ʹ���÷ɻ���Ǩ����������Զ��Ǩ��·�ߵĵ����ڣ��ͷ�����֮���Կɷ���ԭ�ܵء�����ʹ�����“���྿������ʲô������”�����⡣����Ұ��۲졢��־���״�̽�⡢��ҹ��Զ�������Լ����ָ���������ʵ�飬��������ټ�˵����������̽���Σ���δ��ÿ϶����ۡ�Ŀǰ�Ƚ����еĿ����У�
����1.ѵ���ͼ��� ��Ϊ�������һ�ֹ��еġ����Ŵ��������ķ���У�an innate sense of di-rection�������ַ���У����������������Ǩ��Ĺ����У����ϵؼ�ǿ��Ǩ��·�ߵļ��䡣
����2.�Ӿ�����visual orientation������������Ǩ��;���ĵ��Ρ����ۣ�����ɽ���������������Լ���Į�ȣ���Ϊ�쵼�������ϴ�������ᴫͳ��Ǩ��·�ߡ�ʵ��������Ӿ������������̾���Ĺ鳲�����ܲ�����Ҫ�ġ������ڸ�����װ�Զ�Ӵ�����Ȼ���ھೲ15km��130km���ŷɣ������ܰ�ʱ�鳲���ɼ�һ���������Ӿ�����Ķ�����ơ�
����3.���嵼����celestial navigation���ܶ�ʵ�����������������̫�����dz���λ�ö����dz��������ҹ��Ǩ���������Ϊ��Ҫ��
��������̫����λ��ʵ�飬���������ǿ���Ĭ��Kramer��1957��1961������������о����Ѿ���Ǩ��ϰ�Ե�����������д������ڣ��Լ��ش���ʹ�����Ǩ��״̬����ɼ������һ���ķ�����Ǩ�㷽��������ͼ19—55��������������Ϊ�����첻���֡�����þ��Ӵ���̫���ķ�λ������������ɰ�����Ԥ���ķ����������������Զ�볲����ããѩԭ���ͷ�֮������������ֱ������ԭ��ס�أ������������ߣ�һٹ���磬������Ѱ����ȷ��λ����̫����λ���棬������Ϊ�������̫���ķ�λ�ǣ�sun azimuth����ȷ������������Ϊ�����ǽ赱������ʱ̫���Ķ��ո߶������������ԭ��ס�ص�������ԱȽϣ����ⶨ���ڵ�γ�ȡ�������Ϊ������ķ��꣬�ٸ����dz�������ȼ���У������Щ�����д������֤�ļ�˵��
�����dz���������������Sauer��1957���״���Բ�����ڶ�ŷ��έݺ����ʵ��ó��ģ�֤�������ܸ���ҹ�����dz���λ�ö��˺������˴���ʵ���о��������øı������dz�λ�õķ�����Ҳ������������̫�������ʵ��һ����ʹ���ఴԤ���ķ���ı���“Ǩ��”����
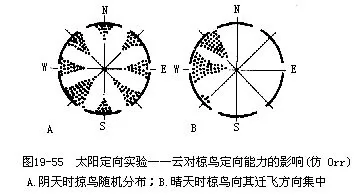
��
������̫�����dz���Ϊ���е�ָ��ʱ�����ڵ������ת���л���������һ���ڲ���“������”��biological clock�����Բ��ϵ���̫�����dz�����Ǩ����֮��ĽǶȡ�������Щʵ��֤���������˹����ոı����—��ҹ���ڣ���ʹ��������������“������”�����µ����ˣ������Ѹ����������������չ�����������ʱ���������ʱ��ͷ���Ǩ�ɡ��ⷽ��Ĺ������д��ڼ��������о���
����4.�Ŷ���magnetic orientation����Ϊ�����ܽ�شŸ�Ӧ��ȷ��Ǩ�㷽�����ѧ˵�������ý�������������Ϊ���ڵ����ڣ���ʹ����ɥʧ����Ķ����Զ�������ģ�Fromme��1961�����Ժ�����д�����ʵ�飬�ر��ǵ¹�����ѧ�ң�Merkel��Wiltschko��1965��Wiltschko et al��1971—1981����������עĿ���о��������˽�����������һЩǨ�ɵ����ࣨŷݺ���������������ܲ���ǿ���˹��ų�����Ȧ��helmholta coils������ѡ���Եظı��˹��ų��ķ���ʵ��֤ʵ����ʹ�������ܿ��������dz���ҹ������£�����Ҳ�����˹��ų�����ĸı�������“Ǩ��”�ķ���������Ϊҹ��Ǩ������ķ���ѡ����Ҫ�ǿ��Դų��ĸ�Ӧ����Ǩ�㷽��ı��������dz�λ���йأ�Ҳ����˵���dz�����У�ش����̵Ķ����dz������ǻ��ڴŶ������Ϣ��
�������ˣ�Southern��1972����Ÿ������ͷ��װ���մɴ����������䶨����ܱ����š�ʵ�����״���Ŵ��д�Ƭ�ļҸ룬��ʹ֮ɥʧ“�鳲”��λ���������������䣨microwave radiation������С������ʵ��Ѱ��һ�ּ���Ǩ������Էɻ���в�ķ�����������Ϊ�������ڲ�ͬ�߶ȣ�2000m��7000m�����Ͽգ�����������ȺǨ������ࣨ������Ѽ�ࣩ��ɻ���ײ����ɻ����������¹ʡ�ʵ�����֤ʵ���������ë�ܶ���������Ӧ����Щ���д���һ���о���
���������������������Ρ����ۡ����塢�ų��ȶ���֮�⣬Ŀǰ���д������ϣ���������������������Ƶ���Ƭ��֤����������һ���ĵ��������£�������������������Ҫ�Ǽ����Եķ磩��ѡ��Ǩ�㷽��������������Ǩ�㡣������Щ��ʵ����˵����������̽���Σ����ڳ���ʾ����Ǩ���붨��֮�գ����д��������о���
�������ģ��о�Ǩ������� ��Ǩ����Ƽ���Ǩ��;�����о��������������Ϻ�ʵ���϶�������Ҫ���塣���������ܹ���ʾǨ�㱾�ܵ��γɼ��䷢չ���̣�Ϊ����������Լ��л����뻷��֮��ĸ��ӹ�ϵ�ṩ��Ϊ��������ϡ���ʵ���ϳ���Ϊ��Ч�����úͿ��ƾ������࣬Ϊ������Ȼ��ϵ�ṩ���ۻ����⣬��Ϊ����ѧ�ṩ�˹������о������ֽ����������Ķ���ϵͳ�����������澫ȷ������ij��������˵����Զ�������ﶨ��ϵͳ��
��
���Ľ� ����������Ĺ�ϵ
��
����������ͼ������Ϊ����ͨ���������������������������е���Ҫ�ԡ������ж��溦�Լ������ͺ�������Ұ��������Դ�������漰�ĸ����ԡ�
��������������ʷ���������������������ľ����漰�������������е���Ҫ�ԣ����Dz��Զ����ģ��������������һֱ��Ѱ��������µ�ѱ��Ʒ�֣��������������������������������ۣ����˼Ҽ�����Ѽ���졢���鼦�����ȵ������Ѵ�Ұ��ԭ��ѱ�����������⣬���������������ҹ����㷺������Ϊ�⡢����Ƥ�P���õ�����ȸ���������Լ�“�������”��ʵΪԭ��ŷ���Ǵ�½��ʯ����Alectoris gracea���ݵȣ����ǽ���ʮ�����������¼���Ʒ�֡�̽���оü�ֵ��ѱ����ֳǰ����Ұ�������Ϊ���������й�����ǰ����Ҳ�DZ���������Ұ��������Դ��һ��;������Ҫ�д�������ѧ�������������о������ڼ���Ʒ�ֵĽ�һ��������������ֳ�������Ŵ�ѧ�Һ�ũѧ�ҵ���Ҫ�����������Ұ������������Ĺ�ϵ��Щ���ܡ�
��������������ࣨ�Լ�Ұ�����������������ģ�������ά����������滷�����Լ���̬ϵͳ�ȶ��Ե���Ҫ���ء�����������������Եı��������ѳ�Ϊȫ���ע���ȵ�֮һ��1992�����Ϲ������뷢չ�����ͨ���ˡ���������Թ�Լ�����ҹ��������Լ��֮һ���е��˱����ҹ���������Ե��������е�Ȼ����Ұ�������ʵ���������Ƕ�Ұ������“��”��“��”��ʶ�����һ�������Ե�ת�䡣����ʷ��չ����������������ʶ�����ſ�ѧ�ķ�չ�����ϸ��º���ġ��������ǿ���Ұ���������ͺ�ʱ����Ұ�Ƚ���խ������ֻ�ǿ����������ֱ������������ʳ���ü�ֵ�߲��ߣ��dzԺ��滹�dz�ׯ�ڣ��Ƿ������ȡ������о�������ŷ��֣������ڻش���������ʱҪ�漰�dz����ӵ����أ���Ҫ���д�������Ŀ�ѧ�о������ҵ���һ��������Ϊ��̬ϵͳ�еij�Ա�����Կ���ʱ���ͻ�֪�����Ƕ������ٵ������˽��̫�٣���Щֻ��Ƥë��Ҫ�������ȥ��������̬ϵͳ���ȶ��Ժ���������Ա�����һ����ԭ���������Ұ������ر���Ŀǰ��ȱȫ����ʶ�ľ������Ұ�����Ҫ���Ƶؼ��Ա������ڴ˻����Ͻ��п�ѧ�ġ��������������á����ھֲ�������ʱ�������Σ���Ķ��Ҫ�ڿ�ѧָ���½����ʵ��Ŀ��ơ�“����”��“����”һ�����֣�Ҫ��ȡ��Ϊ������̬�ȡ�
����������Ŀ���������ã���ʹ��̬ϵͳ��������ȶ��ġ���������������ѭ���Ļ����ϣ�Ҫ�����ġ�����ȵ����ö�����Դ��ȡ����Щ�о��ü�ֵ�ġ�ÿ��ͨ����ֳ����������Ⱥ�е�ʣ�ಿ�֡�����������������Ҳ��һ���˷ѡ������ڶ�����Ⱥ�����洦��������С��һ����Ⱥ���������ṩ����̬Ч��;���Ч��֮�⣬���ڿ�ѧ����������ķ�չ������Ҫ���ס�������������Լ���������ѧ����̬ѧ�����ۣ��������ȴ�����ѧ�о��н�ʾ��������������Ⱥ�еõ���֤�ġ������ڳ������еĵ���������ѧ��������������Ĺ��ף�����������֪�ġ����“����”��“����”���ѳ�Ϊ�Ƚ����ҵ�һ�ֹ㷺��Ⱥ���˶���
�����������˵�ֱ��������ϵ��Ҫ�У�
��
һ������IJ�ʳ����
��
������һ���Բ�ʳ���õĹ��۴���������ܲ�ʳũ�ֺ��棬��ʹ����ʳֲ����ʳ������࣬�ڷ�ֳ�ڼ�Ҳ�Ը���Ӫ����ˮ�ֵ����棨�ر����۳�Ŀ�׳棩����ι���������ƺ�����Ⱥ���������������൱�����á����������ݶ������У�����С������Ҳ��ʳ���棬����ڿ����ͳ溦����������ʬ��ͽ��Ͷ������в��Ĵ����ȷ��棬������Ҫ���á����������������ڶ࣬�ֲ��ڶ�����̬�����ڣ��ر��Ƿ��������ϰ��ʹ֮���漯Ⱥ�ƶ��Ļȳ桢����ȵĻ����Բ�ʳ��������������ʳ������Ⱥ�����ɱ���ġ����Դ������Ͽ����ر��Ǵ�������̬ϵͳ����������������ǣ�Ҫ��ʳ���������������ȫ��ı�������������Ĺ�ʶ��
����Ȼ��Ҫ�ж�ÿһ�������ھ��������ʱ���ڵIJ�ʳ���ã�Ҳ�����������ж���洦��ȴ�����Իش�ģ�������д����Ŀ�ѧ�о��������ܼ���ij��һ���ʳ���������ȫ����������ٺ������������������൱�ڱ����˶�����ʳ����ʳ��������ۣ�ʵ�漰����ʳ���������Եij������ж����Ǻ��棬�ж����Dz�ʳ�����棨��棩��ͨ����ʳ֮��Զ���֮��Ĺ�ϵ����Ӱ�죻�����ض�������ʳ���������Ⱥ����������Ҫ������棩��������̬���ر��������ಶʳǰ���������Ⱥ�ܶȡ���ʵ���ж����ಶʳЧ�����洦��С����Ψһ����“ͨ������IJ�ʳ���ã�����Ҫ�������Ⱥ�ܶ��Ƿ��ѱ������ڲ���Ϊ����ˮƽ”��������ķ�ֳDZ�����ۣ������в���ʹ����Ⱥ�ܶȽ��͵�90�����ϣ��������ĸ����ͨ����ֳ��Ѹ�ٵػָ���ԭ��ˮƽ��
����������������������Ѷȣ�����ﵽ��һ��ȵ��о��ɹ����ࡣ���������еIJ��Ͽ�����Ϊ�������ں�����ܶȽϵ�ʱ���н����ԵIJ�ʳ���ã��������ͻ��ֹ����Ĵ������ӳ������ļ���ڡ������������Ҷ���������̬�о����֣��������꾰��Լ��20����65���׳汻��ʳ�����ں������ʱ������ʳ3.5����7.0�����ҹ��㽭��β���˹����ڵĴ�ɽȸ����ë��IJ�ʳ����һ���꾰Ϊ4.71����22.19����������ë�������ݽ�Ϊ0.22��������20��������Ҫ������ʳ��������Ⱥ����������ԶԶ���ں����������������֪ijЩ�����������ʢ����ݿ��Խ���߷�ֳ����������Ⱥ�������������������ȣ���������ġ����˼���������ij�����ľ�Ҷ�������ݣ�����Ⱥ��������8000�������Դ˷�Ӧ��ǿ��������ݺ��Dendroica cas-tanea��������12������Ȼ������Ȼ�粶ʳij�ֺ���IJ�ֹһ�����ࡣ����ʳ�����������ࡢ�ֲ��㣬��Ժ�����������ã��ر�����ά�������꾰�µ���̬ϵͳ���ȶ����棬���൱��Ҫ�ġ�ͬʱҲӦ��ʶ��������ҵ��Ӫ����ͼ����������ȥ���Ƴ溦�Dz���ʵ�ġ����ر������ڴ�����˹��������־Ӷࡢ���͵�һ�������������ܾӵ����౾����ʮ��ϡ�١�������ɭ�ֺ���ķ��ι�����Ӧ�ᳫ�ۺϷ��εIJ��ԣ�������չ�ͳɱ��ĸ�Ч���ж���ѧɱ��������ö���������������ԭ�塢�������ʳ�����������桢ʳ������ȣ��Լ����͵ĺ������ơ��κε�һ�ķ��η�������������ԡ�
����������ʳ������ı��������� ����ʳ������ĸ���ԭ���DZ�������������Ϣ���������ƴ��вж��Ļ�ѧɱ�����ʹ���Լ���ֹ�Ҳ����ԡ�����һ�����ڵ�����Ҫ�㷺��չ�����������������ȫ������ʶ���ҹ���1988��䲼���л�����Ұ�����ﱣ��������1991������ÿ�꿪չ“������”��������Ѿ��յ����൱������Ч����
�������ֵ��������˹�����������ʳ�����࣬Ϊ��Щ�ڶ�Ѩ�������������ṩ����ij�ַ���ǹ��������ѹ㷺���õķ������ر���ȱ���������������бȽ����Ե�Ч�������Ƕ������˹���������ʳ������Ĵ�ʩҲҪ�ʶȣ����������ҳ�����������������������ࡣ������������ʳ��������ֻ�������������ڶ�Ѩ��������ϲ��ѡ���˹�����ģ������ʳ����Դ����������Լ�������������²���������ʳ��������Ⱥ�����������ࡣ�����������о���֤ʵ����ȸ��С�������еİ�����������ȸ�ֲ��ij������ڵ��˹������У�90�����ϱ���ȸ����ռ�������ڳ����ֵ��������˹�������Ҫ��Ӧ������ͨ����Щ����������Լ��������������Ļ���Ӱ�졣“ʮ����ľ����������”��˼�����Ұ������ı��������Ǻ�ǡ���ġ�һЩ����ý����ν��ij���й�ͨ��“����”ʹ�����ڼ�����“������17��”�ı��������Ƕ�������ʶȱ���˽��һ�ָ��F�硣
�����ҹ������Ϊ�����“ѱ��ŷ���”���Ҳ��һ�������˲Ƶġ���Ⱥ����������������ȣ���ѱ���Ļ�ϲȵ��������ȵ�Ȳ����ǵ���ʳ��������ʳ�����ࣻ��Σ�����“����ǧ�ա�����һʱ”�������Ⱥķ���������������������������е��������������ֳ��ŷ�ʱ���ŷ���Ա���ܴﵽ�ĵص㼫Ϊ���ޣ������������ṩ��ʳ��Ҳ�����ġ�“����һ��ͻ���”��ԭ�����������ʳ��������Ȼ�硣��������һ��ѱ���Ӽ����ݶ�����“�������”����ʹ�Ƕ���������ʳ�����࣬Ψһ���еı���������;��ҲӦ������Ȼ�������Ա�����ʹ����ʳ�����ط�����Ϣ�������غķѴ�������������ȥǿ��“��ķ”���Ѱ���������ת��Ϊ����Ȼ����Ҫ�㷺����Ŀ�ѧ�ռ������Լ��Դ�ͳ��ijЩ�����ʶ���б�
�������������ಶʳ��ֲ��ɢ����Ӱ�� ���������ǻ��۵Ĵ�����ֲ���ڷ��ߣ������������̫�����Ļ���������ȡ���ֲ�����ӻ��ʵΪʳ�����࣬������һЩδ�����������������ų�����Щ��������������������һ���ų������Ӹ������ȷ�������������ķ��ƶ���Ϊɢ������֪һЩ�������ϵ�ֲ����Ǿ���������ɢ���ġ���ѻ����ѻ��ijЩ��ľ�������^����ֲ�����ӵ�ϰ�ԣ��ɽ����ټƵ���Ҷ������������������ص���km����������ڣ�������Ϊ������ʷ��ŷ����������չ����Ҫԭ��
��
������������
��
��������������Ҫ����һЩ����Ŀ������Ŀ����Ŀ���a��Ŀ�Լ�һЩ�������Ƕ��ȡ����Ƕ�����Ⱥ���������Ͽ�ġ��м����Լ�Ⱥ���Լ��⡢��Ⱦ��ü�ֵ�ߵ����ࡣ�ڶ��䷱ֳ������Ⱥ������̬���г���о��Ļ����ϣ��������Ի������ľ������档
�����˶�����������������������Ϊ���У��ڹ涨�������������ڶ���������һ�����֣����ҷ�������֤�Լ������ķ��������룬ÿ�������������Ԫ�ơ�Ϊ����Ӧ������Ҫ����ר�ŵ��о��������������̬ѧ����Ⱥ������̬���г��ڵ��о����˹���������ֳ����������绷��������ɽ�ȵȣ��������ͷŵ�Ұ�����֮�á����������һ�ֱ���Ұ��������Դ�¾������������Ļ�������Ҫ�Ĵ�ʩ��Ҳ�ǽ������ת��Ϊ�ߵ�������Ʒ��һ�־�Ӫ��ʽ��
��
������
��
������������ɵ�Σ�����Ǿֲ��ģ���ʱ������Լ����ǵ���ʶ�̶Ⱥ;���������졣�����Ե���ũҵ�������㡢���ġ��������Լ�ȸ��Ŀ�е�ѻ�ơ�ȸ�ơ�����Ƶ����������ʳ�������ʳ���磬�����������ľ�����ȸ����Ҫ��Ȩ���ʧ�Ļ����ϣ�ѡ�����˵ķ������Կ��ơ�“�ˡ�����ʳ”��ì����������ˮƽ�Լ�����ˮƽ�Ƚϵ͵������ʮ�ּ�������ũ������ˮƽ������Լ��������ı仯������ʶ�ϻ������ı�ġ��亦Ϊ��Ҳ�ǿ��ܵġ�ǰ���ᵽ�ķ��������ƹ����Ե���������һ����
����“��ײ”��bird strike���Ƿɻ����������ȺǨ��������ײ���������¹ʣ�ͨ������ں�����������Ϳշ��е�����¡��Դ�60�����������������������ȡ�����������ƽ���֮�������ֻ������ڽ�����������������ѵ��¹��������ӡ�����ڻ�����ѡַ���ر����غ�������ѡַ��ʱҪ�˽�õ�Ǩ����������ࡢ���ּ��ںͷ�Ǩ�����и߶ȵȡ���������֮��ҲӦ������Ļ���ɽ���ȫ�졢ȫ��ļ�⡣Ҫͨ���Ի���������̬�����ĸ����Լ���չһЩ��������ѧ��������ۺϼ���������������
�����������Я��һЩϸ��������ͼ�����ȣ���Щ���ڼ��ݡ����������֮�䴫��������������ʮ�����������й����Ȳ��������ʸߴ�1/3���ϡ�������֪�������йصĴ�Ⱦ����20���֡������չ���༲�������������о�����������֮��Ĵ���;���Լ������ཡ���Ĺ�ϵ����������ı���������Ľ�������ʮ�����еġ�
��
��ϰ��
��
����1.�ܽ�������Ҫ�����Լ������������Ƶ�Ҫ�㡣
����2.�ܽ�������Ӧ�������ʽ���ڸ�������ϵͳ�ϵĽṹ�ص㡣
����3.���������������������Щ���棿
����4.�����3����Ŀ�ڷ�������������Щ��Ҫ����
����5.�ܽ�����ĸ�����̬��Ⱥ������Ӧ��ͬ�Ļ��������ʽ������̬�ṹ������Щ��ͬ��������
����6.ʼ����ʯ�ķ����к����壿���߱���Щ������
����7.ʲô��Ǩ�㣿����˵������ͺ���
����8.���ֳ��Ϊ����Щ����������������ѧ���塣
����9.��������������Ĺ�ϵ��