��
��һ�� ��Ƥ�����ŵ�����
��
������Ƥ����ȫ�������ں����У�����Ϊ����Գƣ��Ҵ����Ϊ�����Գƣ������Ǵ����γɵģ���������Գ����ε����巢չ������
������Ƥ����Ĵ�����ǻ���������ǻ���ֳƳ�ǻ�ң�enterocoel�������γɣ�����ԭ�����ڣ���ԭ���ı������ɶԵ��ң��Ժ������䣬�γ����߲㣬�����ɴ�����ǻ��
�����������Ƥ����Ƥ��ɣ���ƤΪ����ϸ������Ƥ���������֯������㼰���߲��γɵ��ڹ�������Ƥ����Ϊ��ǻ��Ƥ���ڹ����еļ�С�������ࣩ���е��γɹ�Ƭ����һ����ʽ���У������ࡢ��β�༰���ٺ��ࣩ��Ҳ�еĹ�����ȫ���ϳ�һ�����Ŀǣ������ࣩ���ڹ�����ͻ����������γɼ���̣��Ե�Ƥ���ֲܴڣ��ʳƼ�Ƥ���
������Ƥ�������еĽṹ��ˮ��ϵ��water vascular system�����㣨tube foot�������Ǵ�����ǻ��һ�����ػ��γɵ�һϵ�йܵ���ɣ��п����������ͨ����ˮ�ɽ���ѭ����ˮ��ϵ���������ܣ�ring canal�������ܣ�radial canal���Ͳ�ܣ�lateral canal������ͼ12��7������������������Ĺ��㣬�������˶�����������ʳ���ܡ�������ķֲ�����Ƥ����������������Ϊ10�������й���Ĵ����Ʋ�����ambulacrum��������Ĵ����Ƽ䲽����interambulacrum��������������У�ͼ12-15����
������Ƥ����һ���˶��ٻ�������ϵͳ�йٲ�����������壬���巢�����и��͵��׳棨�����׳桢�����׳桢�����׳桢��β�׳桢�����׳桢��״�׳桢�崥���׳�ȣ���
��
�ڶ��� ��������——���̳�
��
�������̳���Asterias��һ����������ֲ����������������̫ƽ����������ࡣ���̳������ڳ�����Ľ��Ҽ�ס��˶���������ʳ�ԡ����̳���Ҫ������Ϊʳ�����˹���ֳ��˫������һ����Σ�������̳�������ǿ���ҹ��Ʋ�����ϰ������Ϊ���Ϻ��̳���Asterias rollestoni Bell����
������һ���ⲿ��̬ ���̳���������״�������̼�����ɣ����߽������ԡ���һ��5��������ʵ��͵������Գƣ�ͼ12��1Aa����
��
������������������ĩ�˵ľ��루�� R��ʾ���ɴ�100mm���ϡ�����ʱ�������£����������ϡ�����ƽ̹������ɫ����������Ϊ�ڣ���Χ��Χ��Ĥ��peristomial membrane��������������һ���Կ�������˲��IJ�������ambulacral groove����ͼ12��1A֮b���������������2�ţ��������Ϊ4�ţ��������ĩ�������̡���������Ե�߶������пɶ��ļ�����������¡�����Ϻ�ɫ���ƣ���ɫ���ޡ����������봦�и��ţ���С������֮��Ϊ�䲽��������һ�䲽������һ��Բ�ζ��С���ɸ�壨madreporite����Ϊ��ˮ����֮�ڡ�����Ļ����������һ����ֳ�ס�����1�����֣������к�ɫ�۵㣨ͼ12��5�������̳��Ĺ���Ϊ�˶����٣����ڹ���������ţ�������ǰ����
�������̳�����ֲڣ������ڹ�������ͻ���ɼ���papilla����̣�spine��������ֲ��в漬��pedicel-laria����Ƥ����papula����ͼ12��1B�����漬��С����3��С��Ƭ���ɣ�һ����Ƭ��basal ossicle����2���Ƭ��blade����ͼ12-1C����ǯ״���ܻ�����������۹�֮����Ƥ������״��Ϊ����Թ�Ƭ��϶���γɡ����Ϊ��Ƥ����Ϊ��ǻĤ��ͼ12��2������ǻҺ������ѭ�����к�������й�Ĺ��ܡ�
�����������ڲ�����
����1.������Ϊһ����״��Ƥ��������һ�㱡�Ľ���Ĥ����Ƥϸ����ɢ������ϸ�����о�ϸ��������Ϊһ���Ļ�Ĥ����Ƥ�ֿ�����Ĥ��Ϊһ�㡣��Ƥ�Ϻ��������֯�ͼ���㡣�����֯����С��Ƭ������������ϵ��һ�𣬳���״������������Ϊ������ά����Ϊ�ݼ���ά������ƽ�������������ظ������߷���������ݼ����Ϸ��������ڲ�Ϊ��ǻ��Ƥ������ë��ͼ12��3�����ڼ��������Ƥ�ڣ�����һϵ�п�϶���еĿ�϶�γ�һ����Χ����Ƥ���Ļ�����ͼ12��2�����ڹ�������ͻ��������γɼ����⸲����Ƥ��
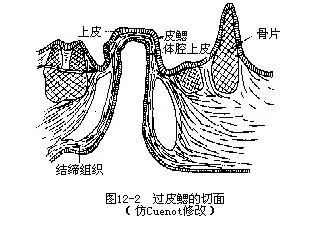
 ��
��
����λ����е��ڹ��������гɴ���״�������������֯������������Ϊ���в������IJ����壨ambulacral plate�������ɲ����ĵױڣ�ǰ����������������С�ף����㼴�ɴ˿�������⡣�����������Ϊһ�вಽ���壨adambulacral plate����������һϸ���Ŀɻ�IJಽ������adam��bulacral spine�����б�������Ĺ��ܡ��ಽ��������Ϊ��Ե�壨supramarginal plate������Ե�壨in��framarginal plate����������������Ϊһ�����ǰ壨carinal ossicle�����������ǰ���Ե��֮��Ϊһϵ�б���壨dorsolateral plate����ͼ 12��4������Щ��Ƭÿһ��˴�������������л�ؽڣ��������������˶���
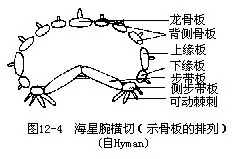
��
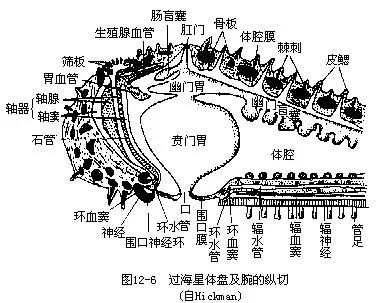
����2.��ǻ ���̳�������ǻ���Χ�������ܺ���ֳ�٣�ֱ������Ķ��ˣ�ͼ12��6������ǻ��һ�����γ���ˮ��ϵ��ΧѪϵͳ��perihaemal system������ǻ�ڳ�����ǻҺ��������ǻ��Ƥϸ������ë�ڶ���ʹ��ǻҺ����������������á���ǻҺ�������ֱ���ϸ��������ǻ��Ƥ���������������ã���ʳ�Ŀ�����Ƥ���ų����⡣�������к��ٵ���ɫС���ε���ǻϸ�����ڡ�
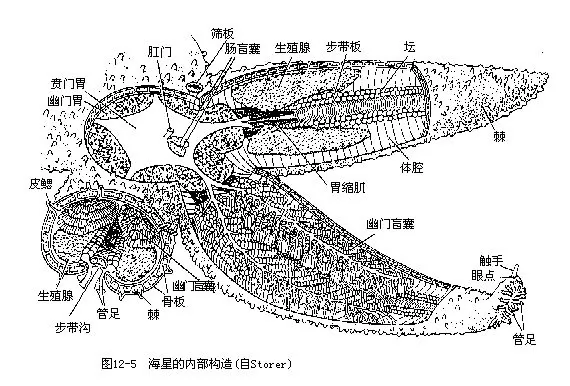
����3.����ϵͳ �����̶ܶ�ֱ���Կ��������档��λ���������У�Χ��Ĥ���봦������Χ����Լ���ͷ��伡��ά�����̵�ʳ���������ij������̵�θ��θ��Ϊ���ڿ��������θ�ͽ��ڷ����������θ�����֣�����֮����һ����������θ����ޣ�����θС����ƽ������������ 2֧ä�ң�������ä�ң�pyloric caeca��������ø��θ��Ϊ�̵ܶij���ĩ�˿���Ϊ���ţ����������ã���������һ��2��3֧�ij�ä�ң�intestinal caeca����ͼ12��5��ͼ12��6����
��
��
���������ܵ���֯����ڣ�ֻ������������ä�ҵ���֯�ṹͬ��������ֻ����Ƥ������ë����Ƥϸ���ɷ�Ϊ��ϸ��������ϸ����ճҺϸ������ϸ������ø������ϸ���ڳ�����֬С�Ρ�������ԭ��ijЩ����——�����ʸ���������B��ʱ�������ʳ�T��ʧ����ä�ҵ���Ƥ���壬����ճҺϸ������ϸ����
�������̳���ʳ�ԣ������嶯���Ƥ�������Ϊʳ���������ţ�������ϴ�Ķ����ʳ������ʱ������������¡�������ס����������ڱ����ϣ��������������ǣ�������θ��������ס��������壬���оֲ���������������θ�ڣ�θ�������ڡ�������Ҫ������θ�н��У���������Ӫ������Ϊ����ä���������棬���ֿ���ä������ǻҺ�ڣ���������������֡�
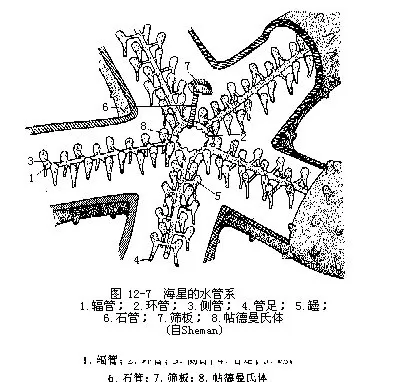
����4.ˮ��ϵͳ Ϊ��Ƥ������������٣��ӷ����Ͽ����Ǵ�����ǻ��һ�����ػ��γɡ�����λ����ڵ���Χ���Դ���ÿ��������һ�����ܣ���������������ܣ���˲����ڹ��㣨ͼ12��7���������ϲ�Ϊһ��״������ampulla��������һ�ܣ�ĩ�˾����̡�����Ϊ���⽻�����У������ÿ����һ���������š�������������ʹ������������ɲ�ʳ����������������Թ���֧���������������ǰ������˶���
��
����������һ�䲽�����������һʯ�ܣ�stone canal����λ�������ڣ����ܱ�����ʯ���ʻ����ʽ�Ӳ����ĩ������ɸ�壬�������ͨ��ɸ��ΪһԲ��С�ǰ壬����������������е�С���ƣ�����������С�ף�Լ200�����������ϼ䲽��������һ�����������壨Tiedmann’ s body����ͼ12��7������ʯ�����ӻ��ܴ�ֻ��һ�������ܹ���9����������������һ�������С�Ͳ������������֯����ǻ������ǻϸ�������ܲ�������ϸ������Щ�����ڴ�λ�������в������ң�Polian vesicle����1��5�����е���ˮ��ϵ�ڵ�ˮѹ���á�
����5.ΧѪϵͳ������ֳgenital sinus���ͻ�ringsinus�������֣�ͼ 12��8����Ϊ������ǻ��һ���֣������ǻ�ҵ�һ���ַ����γɣ��ػ��γɡ���ֳ�λ���������̵�����·���Ϊһ����ιܣ���ÿһ��ֳ�����һ��֧��������γɰ�Χ��ֳ�ٵ��ҡ����λ���棬�ڵ���Χ������֮�¡�ΪһԲ�ιܡ��������һб�и�Ĥ�������Ϊ�ڻ������������ǰ��С�ں��ߡ������������һ����radial sinus����������һ��ֱ��Ĥ��
������axial sinus��Ϊһ���ڹ�״���ң�λ������ɸ��ļ䲽��������ʯ�ܺ����٣�axial gland�������֣����ߵı���ϵĤ��������һ���������ٺϳ�������axial organ����������������СѪ���ɵĺ���״�����֯���ⱻ��ǻ��Ƥ������ڷ���������ֳ���ͨ���˴������һ�������ı��ң�dorsal sac������ǻ������ͻ�롣����ڿ������ڻ�
������Ѫ�ܾ�λ�ڸ�Ѫ��ڣ���˳�ΪΧѪϵͳ��perihaemalsystem����

��
����6.Ѫϵͳ ��ϵͳ���˻���ֻ������Ƭ�Ϸ��ɿ��塣��Ѫ�ܣ�oral haemal ring��λ���ĸ�Ĥ�ڣ��ɴ���������һ��Ѫ�ܣ�radial haemal canal����ͼ12��8�����ڷ��ĸ�Ĥ�д������з�֧�����㡣�Ի�Ѫ���������һѪ�ܴԣ��������ڣ�����λ��ֳ��ڵķ��ڻ�Ѫ�ܣ�aboral haemal ring�����˴��з�֧����ֳ�٣�����ֳѪ�ܣ�λ��ֳ�ķ�֧�ڡ�
����������ͺ������Ѫϵͳ��haemal system�������ԡ�
����7.��������й ���彻����Ҫͨ��Ƥ�����У�����Ҳ����һ�����á���л��������ǻҺ�еı���ϸ����ʳ����Ƥ���ų�����Ҫ�ǰ������ء�
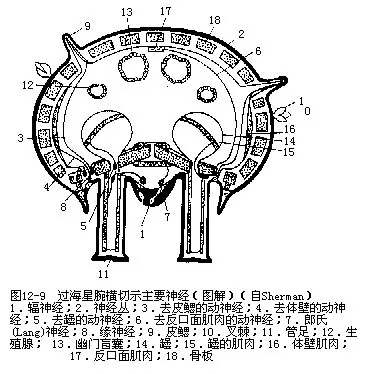
����8.��ϵͳ�о����� ���̳�����ϵͳ����3������ϵ��ϵͳ���ڻ�����ϵ��oral or ectoneural system����Ϊ������������ɣ�ͼ12��9������λ����Ƥ֮�¡���������Σ�λΧ��Ĥ��Χ��������棬����ͨ��Χ��Ĥ��ʳ��������������һ������λ�������ף������֮�£���ֹ�ڶ˴��ֵķ�����һ�࣬�������“V”�Σ����������Ƥ������ͼ12��9������λ�����Ƥ�£�������֮������������ϵ˾�о�������ϵ��hyponeural system��λ���ڵIJ��沿�֣�Ϊһ����5��������ɣ�λ��ǻ��Ƥ�£���֮����������Ƥ��һ��������֯������ϵ����������ϵ��λ������ϵ֮�ϣ��з�֧�����㡢�漬�����ٵļ��⣬˾�˶���������ϵ��aboral system��λ�����棬����ǻ��Ƥ�������������⡣����ϵ������ֻ��5�������ں�������������ٺ������ԣ�˾�˶���
�������̳��ĸо����ٲ��������Ƥ��ɢ������������ε��о�ϸ����neurosensory cell���������д������ͻ�ѧ�о������ֹ��ܣ��ڹ�������̴���Ŀ��࣬�ڼ��Ͳ漬��������Ƥ����ÿmm2�ɴ�70 000�����ڸ���Ķ��˴��ֵĻ���������һ�۵㣬��һȺ�й�ϸ����ɫ��ϸ�����ɣ��ɸй⡣���̳�����Χ�Ĺ�����������ܡ�
��
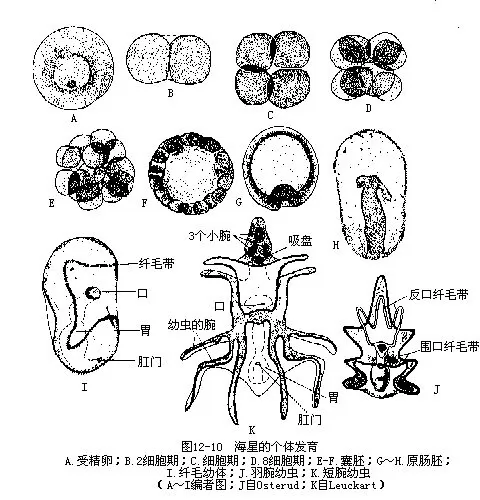
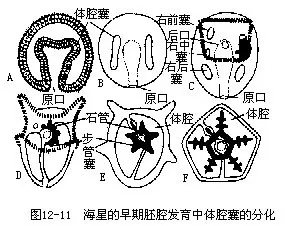
����9.��ֳ���巢�����̳��ڸ����ڻ������෴����䲽����������5����ֳ�٣��������ڽ����������丽�Ŵ���Ϊ��ֳ�ף�ҲΪ5�ԣ���С����ֳ���̡ܺܶ�����ֳ�����ڣ���ֳ�ٲ��������ֳ����ʱ���ɳ������ڣ�������ˣ�ͼ12��5��ͼ12��9�������̳��������壬�����ں�ˮ���ܾ�������ȫ�������ѣ���ǻ���ߣ������ݷ��γ�ԭ���ߣ�ͼ 12��10 A��H��������ǻ�ҷ��γ����߲����ǻ��������������������ȫ��ͬ����̥�������������ӳ���������ǻ��Ҳ��֮�ӳ����ҷ���Ϊǰ���С��������֡��������������ã��Ҳಿ������˻���ʧ��ǰ�в�����������һ��ͬͨ���������ڱ��ࡣһ�Ժ���ǻ�����ϣ������ɶ������Ĵ�����ǻ�������ǻ�ҵ�һ���ַ��룬�γ�ΧѪϵͳ��������ǻ���ݱ��ˮ��ϵ����ǰ��ǻ�ҷֻ�����ǰ����ǻ�ҵĹ�ͬ���ڳ�ɸ�塣��ǰ��ǻ�Һ�������ǻ������̥���������˻���ʧ��ͼ12��11������̥�ӳ���ԭ�����������Ϊ���ţ����ߵ������γɿڣ����Ǻ�ڶ�������������������γɣ���“U”�Ρ����������ë������ˮ����Ӿ����ʳ������ë���壨ͼ12��10I�����˺������������Բ�ε���������������ë�˻���ֻ������Χ�γ���ë������Ϊ�����׳棨Bipinnaria����ͼ12-10J�������ڱ������������ͻ�𣬽�������׳棨Branchialaria���ڣ�ͼ12��10 K����������Գƣ�ǰ�˾���3��С��һ�����̡������׳��ں�ˮ����Ӿһʱ�ں���ˮ�ף�Ӫ������������̬�ڡ��׳�Ŀڡ�ʳ�ܡ����š������˻���ֻ�������м�һ�η������ˮ��ϵ�ķ�������������ԳƱ�ɷ���Գƣ�ԭ���ߵ�����Ϊ����Ŀ��棬�Ҳ��Ϊ�����棬�˻������������γɡ���̬�������������Ϊ����ԳƵ�С���ǡ�
��
�������̳��к�ǿ��������������������������У���������������������������������������Linckia�����⣩��
���� ��Ƥ����ķ���
��
������Ƥ����Ϊһ���ϵ���Ⱥ��ʼ�ڹ���������ͣ���־���͡�ʯ̿�͡�������ʢ���������Լ6000�֣��������ǡ����Ρ���������β�����ٺϵȣ��ҹ��Ѽ�¼ 300���֡����ݶ�������Ρ����ޱ�����ɸ���λ���Լ�����Ľṹ�ȣ���Ϊ2���ţ�5�٣�����4��ȫΪ��ʯ���ࡣ

��
һ���б����ţ�Pelmatozoa��
��
�������������߱�����������������ϣ����������£����������Ŀǣ���Ҫ��ϵͳ�ڷ����档��������Լ650�֣�ֻ��1�٣���4�پ�Ϊ��ʯ�֡�
������һ�����ٺϸ٣�Crinoidea�����ٺ������������У����ܣ�Ӫ�������һ�������߱����ƺ��ٺ��ࣨstalked crinoids����һ������ޱ���Ϊ������ࣨcomatulids��������������Ϣ���ذ�dz���ҽ��ף��ɸ��������������Ӿ���
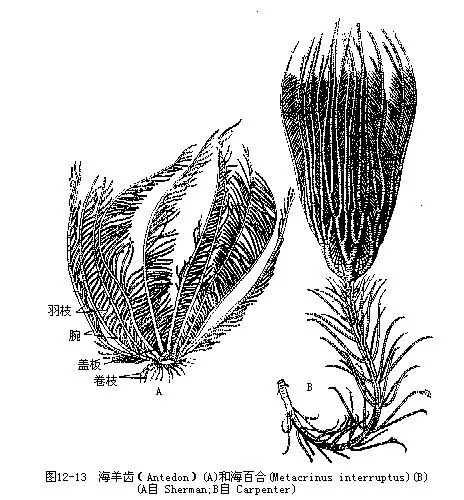
�������ٺ���ָ���������������ͼ12��12������һ��Ʊ���������ǰ幹�ɣ����ϳ��з�֧�ĸ�֧����Ϊ����֧��radiculus�����и������á������ࣨ�����̣����ɣ���ʱ�״��Բ״��������ʯ���ʹǰ���ɣ��߿ڡ����š����������������������֣����˶����ܣ��ɲ�ʳ������ݵ�������̣���ԭʼΪ5����������һ�ٷ�֧���ɶ�������ɶ�����幹�ɣ��������������֧��pinnule������������������Ҫ�Ը��ζ���Ϊʳ����ֳ��λ��ֳ��֧��genital pinnule������֧��һ�֣��ڣ����巢������Ͱ�ε������׳棨doliolaria�������ٺ�������������ǿ���������������һ����䡣��������
�������ٺϣ�Metaerinus���߳�����ͼ 12-13 B������֦��cirrus���࣬�ҹ������зֲ���������ˮ��200m��������ݣ�Antedon����ͼ12��13A���ޱ���Ӫ�����������������ݣ�Compsometra serrata���ൺ�����塢̨���зֲ���
��
�����������ţ�Eleutherzoa��
��
�����ޱ�����������������£���λ�������ǰ�ˣ�����λ����������ˡ�������������Ҫ��ϵͳ�ڿ��档��4�١�
�������������Ǹ٣�Asteroidea�����ƽ����Ϊ�����Գƣ����̺���ֽ粻���ԡ�����ʱ�������£����������ϡ���߲�����������������㡣�ڹ����Ĺǰ��Խ����֯���������Ϳ�����������Ͳ漬��Ϊ������ͻ�𡣴ӹǰ��ͻ����Ĥ����״ͻ���⸲��Ƥ���ڳ���ǻ��Ƥ������ǻ���ڴ�����ǻ����ΪƤ����papula�����к�����ʹ��л������ɢ���������á�ˮ��ϵ������巢���о������׳�Ͷ����׳档����Լ��1600�֡�
��
�����Դ�Ŀ��Phaneroznia��������ϡ���Ե�������������������̣��漬�ޱ���ɰ���ǣ�Luidea�������뷴��������Ե�����Ե������������һ���״��ͼ 12��14 A����
�����м�Ŀ��Spinulosa��������Ե�岻���������������̣������в漬�����ࣨAsterina pectinifera�����ν�����Σ�����߿���״С����̫�����ǣ�Solaster����10������ͼ 12��14 B��C����
����ǯ��Ŀ��Forcipulata��������Ե�岻���������������̣��߲漬Ϊ����ص㡣���̳���Asteriasrollestoni������צ���ǣ�Henricia leviuscula����Բ��ĩ��ϸ��ͼ12��14 D��E������2�־�Ϊ�����غ�ϰ�����ࡣ
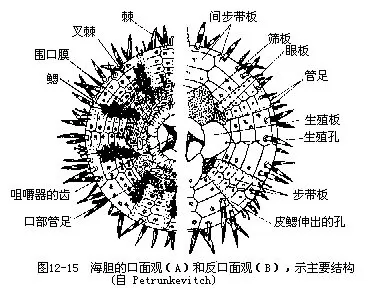
���������������٣�Echinoidea�� ������Σ����λ������Σ������ڹ����������ϣ��γ�һ��̵Ŀǡ��ǰ����������һ�������20�ж���ιǰ壬���г�10������5���߹���IJ�������5������ļ䲽����������������С����ǰ��Ͼ�����ͻ�Ϳɶ��ij������ڶ����ƶ�ϵ��λ���������룬��Χ�ز���periproct����5����ֳ�壬5���۰壨ocular plate����ɡ���ֳ���ϸ���һ��ֳ�ף���һ����ֳ���ף���״���죬����ɸ������á��۰��ϸ���һ�ۿף���ˮ��ĩ���Կ������Ϊ�о�����Χ�ز����и��š�������ΪΧ�ڲ���λ���棬��5�Կڰ壬���й����ڰ�����һ���㡣����Χ��5�Է�֧������Ϊ�������٣�ͼ12��15����

��
���������Ŀ���������ͻ���ɶ���ϸ�������еļ��ܴ֡�����������ھ߽ṹ���ӵľ�����������˹�����ƣ�Aristotle’s lantern�������Ͼ߳ݣ��ɾ�ʳ������ܳ���״�����������ڣ������ࡢˮ����Ϊʳ��
�����������Ϊ�������壬���巢���о������׳棬���̬����������1��2��Ŵ��Գ��졣
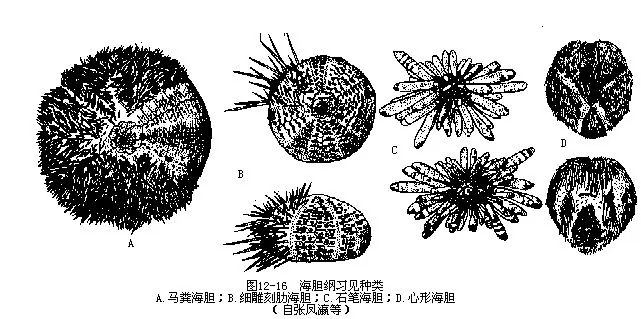
����������800���֣�ϰ�����������ຣ����Hemicentrotus pulcherrimus���ǰ����Σ���ɫ�����̶��࣬״�����ࣻϸ����ߺ�����Temnopleurus toreumaticus���ǽϵ�ƽ�����������ʯ�ʺ�����Heterocentrotus mammillatus�����ִ���ʯ��״��ͼ12-16A��C�����ҹ���ɳȺ���зֲ�����Щ�������Գʰ����Σ�����Գƣ���������Ŀ��Centrechinoidea�������κ�����Echinocardium corda-tum����ͼ12��16D�����������Σ������࣬�ں��ž�λ���棬������Ŀ��Spatangoidea����
 ��
��
�������ģ���β�٣�Ophiuroidea�� ���ƽ����״������С����ϸ�������߷ֽ����ԡ�����������һϵ�����ǣ�vertebra�����Ǽ��пɶ��ؽڣ����ⷢ���ֻ����ˮƽ�����˶����������ϳ��������Ե���Ƭ���������������˻����ʴ���״�����˶����ܡ�ÿһ�����4�������ɣ��������Ҹ�һ�����������������������2�д��ֿף������Դ���������ֿױ��д����ۡ��������˻����������š������ࡢ�п׳桢�л�����мΪʳ��Ҳʳ��ë�ࡢ�����С������巢���о���β���壨ophiopluteus�����������������ͬ�壬̥����
������β��Լ��200�֣����֧��Ϊ����βĿ��Euryalae���������β��Gorgonocephalus�������֧��ɸ�����������϶������������̣�Astrodendrum�����֧�����ƺ�����֧��Ϊ��βĿ��Ophiurae����������β��Ophiura����̲�������㣨Amphiura vadicola�������ɴ�180mm���ϣ��ൺ�зֲ�������β��Ophiothrix fragilis����ͼ12-17��

��
�������壩���θ٣�Holothuroidea�� ������״������Գƣ������Ա⣬�߹��㣬���ೣ�����㣨pa-pillae����һ�ֱ��εĹ��㣩�������̻���̣�warts������λ��ǰ�ˣ���Χ�д��֣�����״����Ŀ�����ͬ���죬����λ��ĩ���ڹ���Ϊ��С��С��Ƭ����״��������������״�������ڻ��ۣ�ĩ������йֳǻ���ɴ˷ֳ�һ�Է�֧����״�ṹ���ƺ�������ˮ�Σ�Ϊ�������еĺ������١��ܴ̼�ʱ���ɴӸ���������ֿ��Ͳ��Ƶк����Ժ������������������ä��״�ľ�ά��������Cuvierian organ��������й���ܡ�Χ��ʳ����ʯ�һ���calcareous ring�����нṹ����5����Ƭ��5�����Ƭ���ɡ�����Ƭǰ���пۣ���ˮ�ܺͷ����ɴ�ͨ����ɸ���˻���λ���ڡ�

��
���������ں������룬ʳ��Ϊ������ɳ�ڵ��л�����Ƭ�����༰ԭ������ȣ���ʳʱ����ͬ��ɳһͬ���롣���巢���о���״���壨auricularia����ͼ12-18���������׳棨doliolaria�����崥���׳棨pentactula������̬���ײΡ�
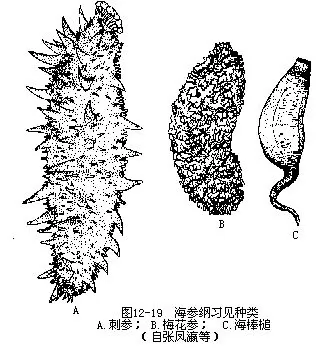
�������δ��غ���������ĺ������зֲ�����Լ1000���֡��ҹ������غ�ϰ�������У��̲Σ�Stichopus japonicus��Ϊ����ʳ�òΣ���ں���̴������ʸߣ�ζ������÷���Σ�Thelenota ananas���峤700mm���ң���Ŀɴ�1m���Ǻ������������࣬������̻�����������÷����״��Ϊ�ҹ��Ϻ�����ʳ�ò�����õ�һ�֡����Ϻ��εĴ��ֲ���֧��ĩ����Сͻ�����J��Ŀ��Aspidochirota��������鳣�Paracaudina chilensis var.ransonnettii���ֳƺ�������ʷĴ��Σ�����ӳ���β״������⻬���������̾��˻��������Ŀ��Molpadonia����ͼ12-19����
��
��
���Ľ� ��Ƥ����ľ�������
��
������Ƥ��������Щ������������棬�����к�������������40���ֿɹ�ʳ�ã����Ǻ������ʸߣ�Ӫ���ḻ�����������̲�Ʒ���ҹ��Ĵ̲Ρ�÷���ε�Ϊ������ʳ�òΡ������ֿ���ҩ������������������ֹѪ֮�����ҹ��ѳɹ��ؽ������˹���ֳ�����̲Σ���������ˮ���о���������ײη���1.5����ɻ���Ϊ55.3�������ý�����������ѿ�ʳ�ã��ݼ����ҹ������������Ժ�����ֳ���ƽ���Ӧ�á�������Ϊ��������ѧ������ʵ����ϡ���������ҩ��������ɢ�ᡢ��̵���ס����ˣ�����������ϡ����Ǽ�����ȸ���Ʒ�������ϣ�������ҩ�������Ƚⶾ��ƽ�κ�θ�����������Ĺ��ܡ��Ժ�������ȡ�Ĵ����նԴ�����ʵ����θ�����н�ǿ���������ã�������Ϊ�о��ܾ���������̥�����ĺò��ϡ���βΪһЩ��ˮ�Եײ��㣨���㣩����Ȼ���ϡ�
��������ϲʳ���壬��Ϊ������ֳ֮������Щ����ļ��ж�������ɶ������Σ��������ϲʳ˫���࣬�ݼ��أ�һ��30���С���ǣ�6���г���50���С���룻һ�����庣��һ����ʳ���ƻ�ĵ�ÿɴ�20������ʺ���Ϊ������ֳ֮�к���
��
����� ��Ƥ�����ϵͳ��չ
��
������Ƥ������ʷ���Գƣ������ǵ��׳�Ϊ����Գƣ���˷���Գ��Ǵ����γɵġ�һЩ�Ѿ����Ļ�ʯ�������е�Ϊ����Գ����Σ�������ں���͵ز��еĺ������ࣨCystidea����ʯ�ͺ����ࣨBlastoidea����ʯ�����������Ϊ��Ƥ���������Ϊ����Գ����εĶԳ��׳棨dipleurula��������3����ǻ�ң�����������ļ�Ƥ�����׳���̬���ơ�Ҳ��һЩ�������崥���׳棨pentactula��Ϊ��Ƥ��������ȡ��崥���׳�ҲΪ����Գ����Σ���3���彺�Һ�Χ�ƿڵ�5���пմ��֣�5���������ܵ���Ϊ��ǻ�ҵ����죬���γ�ˮ��ϵͳ�Ļ������崥���׳����ڽ���Ϊ���������������ת��Ϊ����Գơ���һ�����Ժ���Ӫ����������������Ա����ŷ���Գơ����ٺϸ�Ϊ����ϵ�һ�࣬�����ں���ͣ�������Ժ���˥�䡣���Ǵ����Ӫ�����������̬�����뺣���ܸٺͺ��ٸ����ƣ��Һ��ٺ�����׳��뺣��������ƣ��ʺ��ٺϸٿ�����Դ�ں����ܸ١����Ǹ�����β������һ�£���Ϊ�����Գƣ��������ݻ���ϵ��Ϊ�ӽ���������������β�ٵ��׳��Ϊ�����׳棬�ڽṹ�����ƣ����߹�ϵ�Ͻ���������������Ŀ�����������Σ�����λ���ˣ�����������Գơ��ʺ������ǽ�����β�ٺ�����Գ����εĺ��θ�֮���һ����Ⱥ�����θ�������״������Գƣ��������λ���ǰ�����ˣ��Ǽ�Ƥ�����������һ�ࡣ�������׳��뺣�ٺϸٵ������׳�����ƣ����뺣�ٺϸ����ŽϽ�����Ե��ϵ������ֻ��һ����ֳ�٣����ǽ�Ϊԭʼ����״���ʺ��θٿ������ݻ������н���ķֳ���һ֧��
������Ƥ���ﲻͬ�ڴ����������뼹������һ����ͬ����ڶ��������ǻ�ɳ�ǻ�ҷ����γɣ����߲�����ڹ�������Ҳ�Ǽ�����������������θٵĶ�״������������ﳦ�������ͷ���׳棨tornaria���ڽṹ�Ϸdz����ƣ���˼�Ƥ���������������뼹��������Ϊ���Ƶ���Ⱥ��
��
�������
��
����ë�������Ƽ�����С���峤����40mm���£��������������������Ӫ�������Snadella��Ϊ���ܣ���Ϊ�����и��ζ������ɲ��֣��˶�Ѹ�٣�������Ծ��
����ë�����Ϊͷ�����ɺ�β�����֣������������и�Ĥ�ֿ���ͷ��Բ������Ϊһ���ѵĿڣ�������������ʵĸ�ë���ɰ�����ʳ�����Ĺ��ܡ���С���С�ļ���Ϊʳ�����ɲ���1��2�Բ�����β����һ�����ε�β��������λ���ɲ�ĩ�˸��棨ͼ12-20����
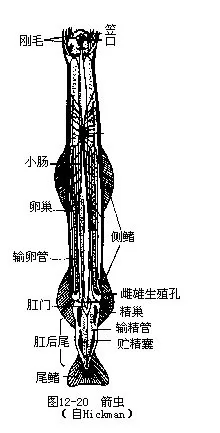
��
������ǻΪ������ǻ����Դ����ǻ�ң��������Ϊͷ�����ɺ�β�����֡���ǻ�ڳ�����ǻҺ�������书�ܡ���ѭ��ϵͳ����йϵͳ��������Ƥ������������������ȫ��ͬ��
����ë���Ϊ����ͬ�壬����������ѳ�һ�ԣ�������ֳ��λ��ĩ�˲��棻����һ�ԣ����쾫������������ų����⡣��Ϊ�����ܾ�����ȫ�������ѣ�����ǻ���ߣ������ݷ��γ�ԭ���ߡ���ԭ����ǰ���γɿڣ�������ڶ�����巢����β���ȷֻ������׳��ڡ�
�����ʱ�����һ���ڣ���Χ����������ǰ����ij����ڣ��γ�һ��������������������һ�����������в��븹���������й�����һ�ԣ�λͷ����
����ë��Լ60���֣��ҹ������Ѽ�¼21�֣����м�������Sag-itta����14�֡����ּ��棨S.enflata��Ϊ�����������֣���ֱ�ֲ�Ϊ0m��250m����99��������100m�ϲ㣬����75����0m��50m�������ֲ���250m��500m�������أ�1985�������������������棨Pterosagitta draco����̫ƽ��棨Krohnitta pacifica���ȣ���ֲ���0m��50m�ϲ㡣��ˮ���й�����棨Eukrohnia hamata����
����ë�����ǻ�γɼ�����ڵ���Դ���ڶ�����ͬ������ṹ��������Ƥ����ǻ�и�Ĥ���ص㣬����Ϊԭʼ��ڶ����м��ػ���һ֧����������ڶ������Ե��ϵ�������С�
��
��ϰ��
��
����1.��Ƥ�����ŵ���Ҫ������ʲô��
����2.ë���ŵ���Ҫ������ʲô��
����3.Ϊʲô˵��Ƥ���ë��Ϊ�������еĸߵ���Ⱥ��
����4.�˽⼬Ƥ�����ë��ľ������塣
����5.������Ƥ�����ϵͳ����������˽���ݻ������塣