第十一章 苔藓动物门(Bryozoa)、腕足动物门 (Brachiopoda)、箒虫动物门(Phoronida)
这三类动物都营固着生活,体柔软,具外壳;身体前端都有由一圈触手构成的触手冠称为总担(lophophore);消化管呈“U”形,肛门位体前方。由于这些共同的特征,过去常将它们隶属于拟软体动物门(Molluscoidea)。这三类动物在系统演化上的类缘关系不清楚,形态结构差异较大,故将它们独立成门,较为妥当。
苔藓动物、腕足类及箒虫三类动物身体不分节,次生体腔;在胚胎发育中,胚孔形成口,这是原口动物的特征;腕足类以体腔囊法形成中胚层及体腔,这又是后口动物的特征。因此这三类动物可能介于具有次生体腔的原口动物和后口动物之间的一类动物。
第一节 苔藓动物门
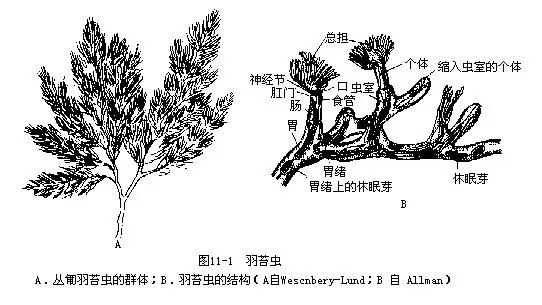
苔藓动物为群体,营固着生活,外形似苔藓植物,故名。大多数生活在温带海域,少数淡水产。群体的每个个体(zooid)很小,不及1mm,外被一由外胚层分泌的角质或钙质的虫室(zooecium)。
个体头部不明显,前端体壁外突,于口周围形成圆形或马蹄形物,其上生有触手,触手具纤毛,这是触手冠,称为总担,为摄食器官。总担可伸缩出入虫室顶端的开口。总担中央为口,消化管呈“U”形,内壁上皮具纤毛。肛门开口于总担的外侧(图11-1B),故又称外肛动物(Ectoproc-ta)。缺乏肾管和循环系统。苔藓动物为雌雄同体,能进行配子生殖。每个个体具有一个在体腔膜上形成的卵巢和发生在体腔膜形成的胃绪(funiculus)上的多个精巢,受精卵在体内或体外发育。海产种类有一似担轮幼虫的幼虫期。通常以出芽法行无性生殖。可产生休眠芽,外被几丁质壳,能抵御不良环境。神经系统不发达,皮下神经网状,神经节在背侧,位口和肛门之间,发出神经至触手等处。
苔藓动物的生存种类约 4000种,化石种有15000种。
被唇纲(Phylactoloemata)总担呈马蹄形,口背侧具一上口突(epistome)(又称上唇,实为口背侧的一体壁突起);淡水产。淡水苔虫北京产3种;杭州有5属8种,羽苔虫属(Plumalella)(图11-1A)中的4种:灌木羽苔虫(P.fruticosa)、丛匐羽苔虫(P.repens)、裂头羽苔虫(P.em-arginata)及斑条透明苔虫(Hyalinella punctata)为世界性种。拟菌羽苔虫(P.fungosa)除杭州外尚分布于北京、沈阳、南京等地。
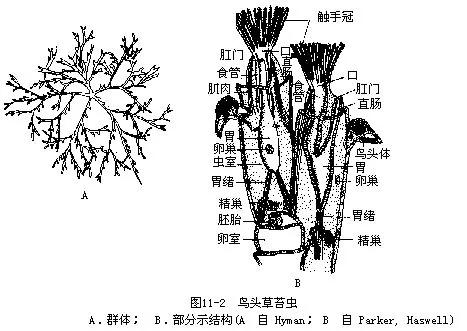
裸唇目(Gymnolaemata):总担为圆形,无上口突;绝大多数海产,我国已报道199种,鸟头草苔虫( Bugula avicularia)(图11-2),除正常个体外,尚有鸟头体(avicularia),形似鸟头,颚可开闭,能除去体上的污物。这是群体的多态现象。棘苔虫(Scrupocellaria)具有鞭器(vibraculum),刚毛状,摆动可除去体上的外物。
海产苔藓虫为海洋污损生物的主要成员之一,其生态特点是在船底及一些设施上形成特定的生物群落。沿海工厂冷却水管、船底、浮标、码头、水产养殖网箱等设施及养殖的海带、贝类等都有苔藓虫群落附着,造成不同程度的危害,阻碍养殖生物的生长发育,使产量下降。我国黄渤海沿岸有苔藓虫35种,加州草苔虫(Bugula californica)在旅顺港、塘沽新港、蓬莱港、烟台港、连云港等都有分布。
第二节 腕足动物门
腕足动物全部生活在海洋中,多数分布在浅海。体外具背腹两壳,很像软体动物,故以前将其归入拟软体动物门,但这两类动物差异极大。
腕足类的背壳小,腹壳大,腹壳后端常具一肉质柄(peduncle),以固着外物(图11-3A、 B)。肉质柄收缩,使动物快速潜入海底泥沙中。背腹二壳内面各具一片外套膜,其边缘有刚毛。壳由套膜分泌形成,二套膜之间为外套腔,动物的柔软身体的大部分位于其中。外套腔被隔膜分为前后两部,前部内有螺旋状的总担,一般左右各一,后部为内脏团(图11-3C)。腕足类体腔发达,充满体腔液;循环系由心脏和血管组成,血管与体腔相通,故循环为开管式,血液即体腔液。总担的基部为口,消化管呈“U”形,有的种类无肛门。由于总担上触手的纤毛摆动,造成水流,摄食粘着的藻类及有机颗粒,送入口中。总担除有摄食功能外,又是腕足类的呼吸器和幼体孵化袋(Ri-chardson,1987)。具1或2对后肾管,兼有生殖导管功能。腕足类雌雄异体,一般具有2对生殖腺。胚胎发育中以肠腔囊法形成中胚层及次生体腔,这是后口动物的特征。个体发生中有似担轮幼虫具纤毛的幼虫期(图11-4)。神经系统不发达,食管周围有一神经环,由此发出神经至体各部。无特殊感觉器官,外套膜边缘触觉灵敏。遇外界刺激,闭壳肌收缩,关闭背腹壳。
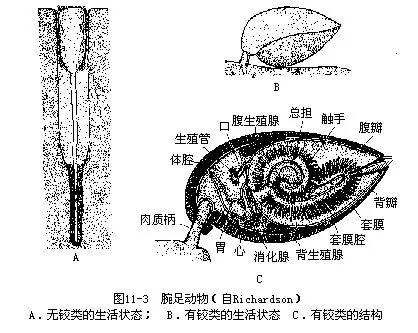
腕足类生存种类有300多种,已描述的化石种在30 000种以上。下寒武纪出现,奥陶纪至二叠纪最繁盛,中生代时大为减少,到新生代时大部分种都绝灭了。腕足类的化石对鉴定地层和石油开采有重要的参考价值。腕足类生活在不同深度的海水中,从潮间带至4000m深海均有分布。
无铰纲(Ecardines)背腹两壳几乎相等,壳多为几丁质;二壳由闭壳肌连在一起;有肛门。我国沿海分布有海豆芽(Lingula anatina)(图11-5),壳长方形,长约25mm,柄长。大海豆芽(L.murphiana)的壳长60mm~70mm,数量较少。烟台、大连、青岛及北戴河均有分布。
有铰纲(Testicardines)背壳小,腹壳大,多为钙质;二壳由齿和槽绞合;无肛门。酸酱贝(Terebratella coreanina),柄短。南极海域有有铰类动物分布(Richardson,1986)。
第三节 箒虫动物门
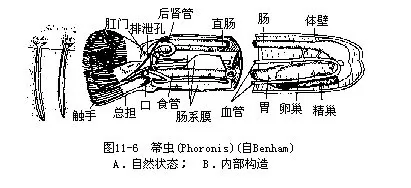
箒虫体呈蠕虫状,长6mm(Phoronis ovalis)至200mm(Phoronopsis viridis),大多数不超过100mm。管栖,管子由上皮分泌,成分为几丁质。箒虫全部生活在浅海海底泥沙中,上端外露(图11-6A)。
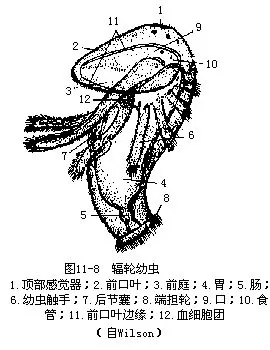
箒虫体前端具一马蹄形总担,由内外两行具纤毛的触手构成,围绕着口。口为横裂状,位两列触手之间;消化管“U”形,肛门在总担基部,口的一侧。次生体腔,被一稍斜行的隔膜分为前后两部,前部为体腔,后部为后腔(metacoel),后腔又为背、腹、侧肠系膜隔成4个纵室。闭管式循环,无心脏,背、腹血管可以收缩。红血细胞含有血红蛋白。具后肾管一对,“U”形,兼作生殖导管用。肾孔开口于肛门附近(图11—6B)。多数种类雌雄同体,少数雌雄异体。雌雄同体种类的卵巢位侧血管的背侧,精巢位腹侧。卵裂有各种形式,有的为螺旋式卵裂,个体发生中经一似担轮幼虫的辐轮幼虫(actinotrocha)(图11-8)。神经系统简单,口后有一上皮内神经环,由此发出神经至身体各部。
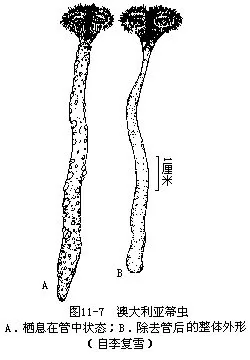
箒虫种类很少,只有2属20余种。分布仅限于热带和温带的浅海区域。澳大利亚箒虫(Phoronis austrulis)(图11-7)在厦门鼓浪屿发现,体最长者为93mm。此种分布于澳大利亚、日本及印度等。
第四节 苔藓动物、腕足动物及箒虫动物的系统发展
这三类动物的演化地位较难确定。它们在个体发生中具有似担轮幼虫的幼虫阶段;都为次生体腔;后肾管;生殖细胞来自体腔上皮,由肾管排出;这些特点似环节动物。腕足类以肠腔囊法形成中胚层和体腔,这是后口动物的特点。因此这三类动物可能是介于原口动物和后口动物之间的一类动物。尽管它们均为次生体腔,具总担,“U”形消化管,有似担轮幼虫的幼虫期等特征,但苔藓动物有虫室,除有性生殖外,尚进行无性生殖(出芽);腕足类具背腹2瓣壳;箒虫体呈蠕虫状,体腔有隔膜,闭管式循环系统。这些都说明它们在系统演化上彼此的类缘关系不密切。
复习题
1.苔藓动物门、腕足动物门及箒虫动物门各有何主要特征?
2.为何过去将苔藓动物、腕足动物和箒虫动物三类归属于拟软体动物门,其依据是什么?三类动物各具何结构特点而独立成门。
3.试述这三类动物介于原口动物与后口动物之间类型的理论依据