第一节 概述
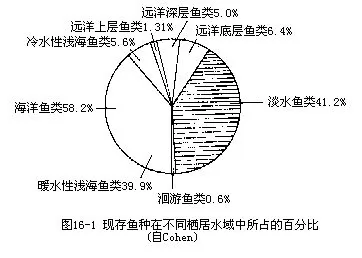
鱼纲是体被骨鳞、以鳃呼吸、用鳍作为运动器官和凭上下颌摄食的变温水生脊椎动物。除极少数地区外,不论从两极到赤道,或是由海拔6000m的高原山溪到洋面以下的万m深海,都有鱼类生存。它们在长期的进化过程中,经历了辐射适应阶段,演变成种类繁多、千姿百态、色彩绚丽和生活方式迥异的22000多种鱼类。鱼纲是脊椎动物中种类最多的一个类群,超过其他各纲脊椎动物种数的总和,包括硬骨鱼和软骨鱼两大类。生活在海洋里的鱼类约占全部总数的58.2%,栖于淡水中的鱼类约占41.2%(图16—1),显然这一现状与海洋的面积辽阔及环境条件比较复杂有关。1954年于南海广东省沿岸捕获的鲸鲨(Rhineodon typus)就是世界上最大的鱼种,其长度可达20m,重量超过5t。最小的鱼是生活在菲律宾淡水湖内的邦达克虎鱼(Penda-ka Pagmaeae)成鱼体长仅12mm,不及鲸鲨的1600分之一,也是世界上最小的脊椎动物。鱼类生存的水温适应幅度较广,既有栖息于52℃山间温泉的花鳉(Cyprinodon macularins),也有可以忍受北极地区水温在-2℃以下的黑鱼(Dallia pectoralis)。
鱼类出现于古生代的志留纪,到泥盆纪已演化出四大类:棘鱼类(Acanthodii)、盾皮鱼类(Placoderma)、软骨鱼类(Chondrichthyes)和硬骨鱼类(Osteichthyes)(图16-2)。

棘鱼类是原始有颌动物,体表覆盖一层“细密”的菱形鳞片,头侧有骨质鳃盖,奇鳍前方有棘一枚,2对偶鳍之间尚有5对小棘,代表动物为栅鱼(或称梯棘鱼)。盾皮鱼类体被盾甲,具偶鳍、歪形尾和软骨性骨骼;在石炭纪绝灭。最早发现的古软骨鱼类化石是裂口鲨(Cladoselache),已具有盾鳞、歪尾等许多现代软骨鱼类的特征。最古老的硬骨鱼是古鳕类(Palaeoniscoidea),由此演化出古内鼻孔鱼类(Choanichthyes)和现代硬骨鱼类主体辐鳍鱼类(Actinopterygii)。鱼类的发展经历了泥盆纪的初生时代、中生代的中兴时代,到新生代达到全盛时代,成为脊椎动物中的最大类群。
第二节 鱼纲的主要特征
水是鱼类的唯一生存环境,离水后将无法存活,并因鳃的粘连和表面干燥,造成窒息而很快死亡。只有极少数鱼类,由于具有特殊的适应器官及机能,才得以在离水后的短时期内苟延残喘。鱼之所以能在水中生活,主要决定于水的一系列理化特性和鱼类具有适应水生环境的形态特征及其生理机能。
水的密度和比重远远大于空气,对动物在水中运动会产生较大的阻力,但是密度较大的水体却又能轻而易举地使鱼体飘浮起来,不需有力的四肢支撑体重,这是水和陆地2种生活环境的主要区别,也是水能成为鱼类生活环境的重要特征之一。水体由含有巨大热量的介质构成,热容量大,因此水温的变化辐度较小,而海洋的温度几乎趋于恒定。此外,水还有大量的汽化潜热,每当水体温度出现周期性变化时,可产生表层冷水下沉的垂直对流现象,并在不断降温的情况下,水面冻结成冰层,冰的比重较小,能浮于水面,水的这种性质不但可以通过垂直对流,为深层和底牺性鱼类供给营养物质及氧气,也能有效地阻止冰层下的水温继续降低和散热,有利于鱼类生活和越冬。
一、体形和皮肤
(一)体形 鱼类由于生活习性和栖息环境不同,分化成各种不同的体型。多数鱼类生活在温带和热带海洋水深200m内的中上层,具有纺锤形(fusiform)的体型,能作快速而持久的游泳。栖息在江湖河池和静水水域中的鱼类,一般都有与纺锤型相似的侧扁型(compressiform)体型,这些鱼类游速较慢,不太敏捷,很少作长距离迁移。还有适应底栖生活的平扁型(depressiform)体型以及潜伏于泥沙而适于穴居或擅长在水底礁石岩缝间穿绕游泳的鳗型(anguilliform)体型等。
鱼体可分为头,躯干和尾三部分。头和躯干之间以鳃盖后缘(或最后一对鳃裂)的鳃孔为界,而躯干与尾的分界线是肛门或泄殖孔。鱼类不仅具有背鳍、臀鳍和尾鳍等奇鳍,还出现了偶鳍(包括胸鳍和腹鳍)。鳍膜内有鳍条支持,鳍条包括棘(spine)和软鳍条(soft ray)两类,软鳍条又可分为分节而末端分支的分支鳍条(branched ray)和分节而末端不分支鳍条。棘和软鳍条的数目依种而异,是鱼类分类学上的鉴别特征之一。胸鳍位于头的后方,是协助平衡鱼体和控制运动方向的器官。鳐、等软骨鱼类的胸鳍扩大,与躯干结合成盘状;马鲅(Eleutheronema)、鲚鱼(Coilia)、红娘鱼(Lepidotrigla)、鬼鲉(Inimicus)的部分胸鳍鳍条呈游离的长丝状或指状;海鳝、黄鳝(Monopterus)和舌鳎等少数鱼类无胸鳍。腹鳍具有稳定身体和辅助升降的作用,通常体积小于胸鳍。许多比较低等的鱼类(鲱形目、鲤形目、鲇形目等)腹鳍都位于腹部;鲈形目、金眼鲷目等鱼类的腹鳍位于胸鳍下方前后,称腹鳍胸位,虎鱼类和狮子鱼类的左、右胸鳍常于此连合在一起,形成杯盘状的吸盘,能吸附在岩石上不被水流冲走。有些鱼类(鳚类、鼬鳚类)的腹鳍位置可前移至喉部,称腹鳍喉位。缺乏腹鳍的鱼类有鲀、鳗鲡、黄鳝、海龙、带鱼和箭鱼等。背鳍位于背部正中,它的形状、大小和数目因鱼而异。高等的棘鳍鱼类(Acanthopterygii)有2个背鳍,第一背鳍由棘组成,第二背鳍主要由软鳍条组成;低等的软鳍鱼类(Malacopterygii)只有一个由软鳍条组成的背鳍,有些种类(鲑、大麻哈鱼、黄颡鱼等)在单个背鳍之后还有一个脂肪性的脂鳍(adipose fin),鳕鱼类有3个背鳍则是鱼类中的罕见例子。臀鳍(anal fin)位于肛门和尾鳍之间,是维持鱼体垂直的平衡器官。尾鳍在鱼的运动中起着舵和推进作用,除了鲼、海马、黄鳝、蛇鳗的尾无尾鳍外,大多数鱼类的尾鳍都是在外观上呈上、下叶对称的正尾(homocercal),只有鲨、鳐和鲟鱼等具有上叶小、下叶大的歪尾(heterocercal)。有些种类如鲼、海马和黄鳝的尾鳍已退化消失。
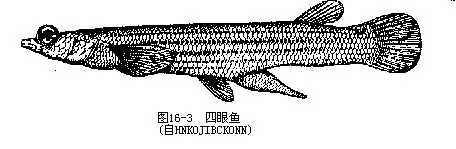
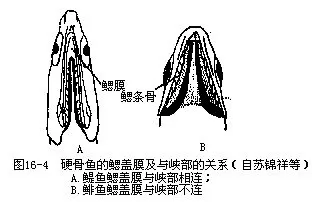
口位于头的前部,由活动性的上、下颌支持,这是脊椎动物中从鱼类开始出现的结构,因此,鱼纲、两栖纲、爬行纲、鸟纲和哺乳纲动物合称为颌口类(Gnathostomes)。颌的出现在脊椎动物发展史上,是一个极其重要的形态发展和进步,并由此引起生活方式的重大改变:动物可以用上、下颌构成的口作为索食工具,主动地追逐捕食对象,增加获取食物的机遇,并通过口中牙齿的撕咬和压研作用,使原来不能直接利用的物质转变为食物,从而开拓了广泛摄取食源的领域。上、下颌既是鱼类索食、攻击和防御器官,也是营巢、求偶、钻洞和呼吸进水时的工具。颌的出现及其多用途的活动机能,还促进了运动器官、感觉器官和其他相关器官的发展,从而带动了动物体制结构的全面进化。有些鱼类在口的周围长有1~5对触须(barbels),触须上分布有味蕾,司味觉功能。根据触须的着生部位不同而分别命名为吻须、颌须和颏须等。眼一对,侧生,眼的大小和位置随各种鱼类的体型及生活方式而异,比较特殊的有鲽形目,为双目并列的“比目鱼”;在黑暗无光的深渊或岩洞地下水中生活的鱼类(我国云南的盲条鳅Nemacheilus gejiuensis和裸腹盲鲃Typhlobarbus mudiventris等),眼被皮肤所覆盖,逐渐退化成盲鱼;生活在美洲河川水面的四眼鱼(Anableps tetraphthalmus)具有适于观察水陆2种环境内物体的眼睛,眼的上、下半部功能不同,其内部构造也发生了相应的变化(图16—3)。鼻孔一对,大多位于吻的背面,是嗅觉器官的通道,每个鼻孔由瓣膜分隔为前鼻孔和后鼻孔,水流通过鼻孔进入鼻腔与嗅囊发生接触,能感受外界的化学刺激。除肺鱼类和总鳍鱼类外,鱼类的鼻腔均不与口腔相通,但虎鲨和须鲨(Orec-tolobus)等有连接鼻孔及口角的鼻口沟(naso-oral groove),被认为是外鼻孔通口腔的最原始通道。头的后侧有一骨质鳃盖(opercular),后缘内侧附生鳃盖膜(gill membrane),鳃盖下方为容纳鳃的鳃腔(branchial cavi-ty),其内、外分别与咽部及鳃孔相通,进入口中的水流通过鳃腔由鳃孔排出体外。鳃盖膜在头部腹面是否与峡部(isthmus)相连,是淡水鱼类分类的鉴别依据之一(图16—4)。鲨鱼和鳐鱼等软骨鱼类无鳃盖及鳃腔,咽部有5~7对鳃孔,直接开口于体外。躯干两侧各有一条与背部轮廓大体平行的侧线,由皮内侧线管开口在体表侧线鳞上的小孔连接而成。此外在头部也常常形成复杂的侧线系统。有些鱼类的侧线可多达3条(舌鳎)至5条(六线鱼),而鳑鱼、短尾(Phoxinus brachyu-rus)、花江(P.czejanowskii)、中华细鲫等侧线不完整或完全消失(林氏细鲫Aphycypris lini)。
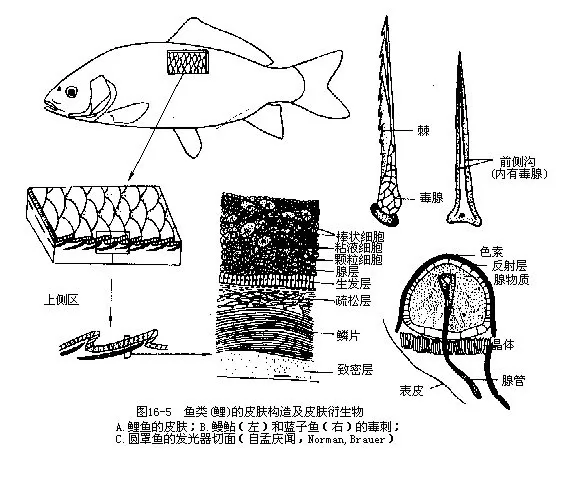
(二)皮肤及其衍生物 鱼类的皮肤由表皮和真皮组成,还有色素细胞、毒腺、发光器和鳞片等皮肤衍生物及附属结构。皮肤的功能主要是用于保护身体,但有些鱼类尚有辅助呼吸、感受外界刺激和吸收少量营养物质的机能。表皮可分为基部的生发层和上部的腺层,生发层细胞具旺盛的分生能力,是产生新细胞的增殖层;腺层因内含杯状细胞、颗粒细胞、浆液细胞、棒状细胞、线细胞等单细胞腺而得名。单细胞腺分泌的大量粘液,能滑润鱼的体表,减少游泳时与水的摩擦,鱼体只需消耗较少的能量,即可获得较快的运动速度。粘液还能保护鱼体使之免遭病菌、寄生物和病毒的侵袭。粘液有迅速凝结和沉淀水中悬浮物质(泥沙、污物)的作用,这对栖息在水质混浊环境中的鱼类说来,更具有特殊的意义。真皮的厚度大于表皮,位于表皮层下方,由纵横交错的纤维结缔组织(胶原纤维和弹性纤维)组成,自表及里可分为外膜层(membrana externus)、疏松层(stratum spongiosum)和致密层(stratum compectum)等 3层。真皮层下面有一层不太发达的皮下层(subcutis),内含色素细胞、脂肪细胞和供应皮肤营养的毛细血管等。色素细胞(chromato-phore)有4种,即黑色素细胞、红色素细胞、黄色素细胞和虹彩细胞(或称反光体),丰富多彩的鱼类体色就是由于各种色素细胞互相配合而成。毒腺(venomous gland)由许多表皮细胞集合在一起,沉入真皮层内,外包结缔组织,特化成一个能分泌有毒物质的腺体。毒腺与刺、棘的关系比较密切,常位于牙(海鳝)或刺棘的基部及其周围(、鲼、角鲨、虎鲨、毒鲉、腃、黄颡鱼等),毒液可通过棘沟或棘管注入其他动物体内,达到自卫、攻击或捕食的目的。人们在捕鱼时如不慎被毒鱼咬伤或刺伤,会发生伤口剧痛、麻木、出血、肿胀现象,严重者能并发恶心、呕吐、盗汗和呼吸促迫等症状,可导致组织腐烂、肢体麻痹、心力衰竭而死。许多深海鱼类的体表具有适于在黑暗环境生活的发光器(luminous organ),其形状、大小、数目、位置及构造随鱼种而不同。典型的发光器一般由下部的发光腺、上部的晶体及包裹在外面的反射层。色素罩等各部组成。发光腺可分泌一种含磷的荧光素,在荧光酶的作用下,能被血液中的氧所氧化,成为氧化荧光素而发出不同颜色的冷光,用于照明、寻觅食物或识别同类。氧化荧光素只有在氯化钠溶液以及与海水等渗的各种盐类溶液中才能发光(图16—5)。
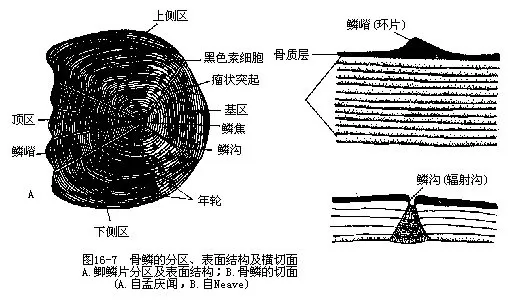
大多数鱼类的全身或一部分被有鳞片,具有保护作用,只有少数鱼类无鳞或少鳞。鱼鳞分3种,即骨鳞(bony scale)、盾鳞(placoid scale)和硬鳞(ganoid scale),分别被覆于硬骨鱼类、软骨鱼类及硬鳞鱼类的体表(图16—6)。

骨鳞是鱼鳞中最常见的一种,是真皮层的产物,仅见于硬骨鱼类。骨鳞柔软扁薄,富有弹性,表面可分为基区(前区)、顶区(后区)、上侧区和下侧区,在偏近前区处有一鳞焦(scale focus),是鳞片的最早形成部分。基区斜埋在真皮的鳞袋内,前后相邻的鳞片作覆瓦状排列于表皮下,顶区露出部分的边缘因呈现圆滑或带有齿突而被称为圆鳞(cycloid scale)及栉鳞(ctenoid scale)。骨鳞分为上、下2层,上层为骨质层(也称骨片层或透明齿质层),脆薄而坚固,表面有环圈状的隆起线,叫作鳞嵴(scale ridges);下层为纤维层(也称纤维板或基板),由成层的辐射纤维和环状纤维排列而成。上、下两层的生长方式不同,上层是从原来的骨质层边缘向外逐圈添加,中央部和外周部的厚度始终是几乎相等的;下层是一片一片地从底部中心往外缘生长,即每次新长出的一片总是重叠在最底层,而且比老的上一层长得大一些,因此鳞片的最厚处总是位于中央部,并将随同鱼鳞增大逐年加厚。鱼鳞数目终生不变,但能继续增大,可用作分类鉴定特征之一。鱼鳞表面的鳞嵴间距随生长强度而变化,这是外界环境影响及鱼体内营养物质摄取状况在鳞片上的反映,在冬季生长缓慢时期,鳞嵴显得微弱而狭窄,相互接近,甚至出现中断、改变走向、波曲等情况,当春夏之际进入生长恢复期时,在缓慢生长区的鳞嵴边缘产生许多新的、连续的和间隔宽阔的环形鳞嵴,鱼体周期性有规律的生长及其在鳞片表面留下的鳞嵴变化痕迹,每年形成一个宽、窄相间的生长带,即为年轮,可用作确定或估计鱼龄的标志(图16-7)。骨鳞可发生各式各样的变异,形成鲹科鱼类侧线上的骨质棱鳞、刺鱼体侧的骨板、包在玻甲鱼(Centriscus scutatus)体外的透明骨甲和箱鲀的骨箱等,以适应不同环境和特殊的生活方式。
盾鳞为软骨鱼所特有,平铺于体表互成对角线排列,可使流经表面的水流流态平顺,涡漩减少,有助于提高游泳速度。盾鳞由菱形的基板和附生在基板上的鳞棘组成,棘外覆有釉质,其构造与牙齿相似,血管、神经可穿过基板孔进入鳞棘的髓腔内。
硬鳞只存在于少数硬骨鱼的硬鳞鱼类(鲟、鳇、弓鳍鱼、雀鳝、多鳍鱼等),来源于真皮层,鳞质坚硬,成行排列而不呈覆瓦状,在一定程度上影响了鱼体活动的灵活性。分布在我国的鲟鱼和鳇鱼除了尾鳍上叶保留着若干硬鳞外,其余的均已消失不见,至于体侧5行块状骨片是骨板而非硬鳞。
二、骨骼系统
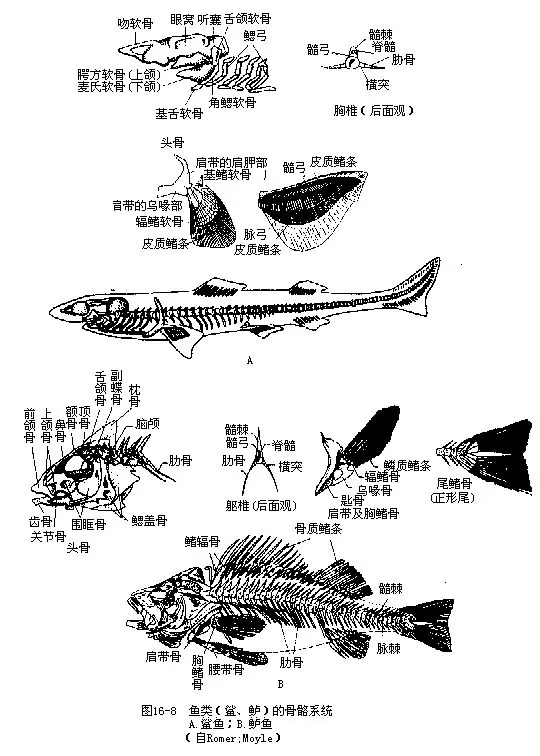
鱼类已具较发达的内骨骼系统,按其功能和所在部位,以及胸、腹鳍的出现,可分为中轴骨骼和附肢骨骼两部分。中轴骨骼包括头骨和脊柱,附肢骨骼包括带骨和鳍骨(图16-8)。鱼类的骨骼系统由软骨或硬骨组成。软骨质的骨骼系统是圆口纲动物和软骨鱼类的基本特征。硬骨鱼类的骨骼系统是由两种不同发育途径形成的硬骨所构成,即从软骨骨化而成的软骨化骨(cartilagobone)和在膜质基础上直接骨化而来的膜骨(membrane bone),二者在形态结构及化学组成上都很难区分。
(一)中轴骨骼(axial skeleton)
1.头骨(skull)可分为包藏脑及视、听、嗅等感觉器官的脑颅(neurocranium)和左右两边包合消化管前段的咽颅(splanchnocranium或visceral skeleton)二部分。鱼类具有完整的脑颅,由一块箱状的软骨或许多骨片拼接而成,只留有口、鼻孔、眼和鳃孔等器官未被包在脑颅内。构成脑颅的骨块数多于脊椎动物中的任何一纲,这些骨块分别位于脑颅的鼻区(olfactory region)、蝶区(orbital region)、耳区(otic region)、枕区(occipital region),以及脑颅的背、腹和侧面。鼻区有中筛骨(mesethmoid)和侧筛骨(ectoethmoid);蝶区有组成眼窝前、后壁的眶蝶骨(orbitosphenoid)和翼蝶骨(alisphenoid),脑颅侧面围绕眼眶四周的是数目不等的围眶骨(circumorbital);耳区前接蝶区而包围耳囊,本区有蝶耳骨(sphenotic)、前耳骨(protic)、翼耳骨(pterotic)、上耳骨(epiotic)和后耳骨(opithotic)等;枕区是脑颅的最后部,由围绕枕骨大孔的上枕骨(supraoccipital)、侧枕骨(exoc-cipital)和基枕骨(basioccipital)组成。此外,脑颅背面自前往后还覆有成对的鼻骨(nasal)、额骨(frontal)、顶骨(parietal)、颞骨(temporal),腹面则有犁骨(vomer)和副蝶骨(parasphenoid)各一块。
咽颅是7对分节的弧形软骨,位于脑颅下方并围绕着消化管的前段。第一对为颌弓(man-dibular arch),在软骨鱼类中构成上、下颌,这是脊椎动物最早出现和原始型的颌,称作初生颌(primary jaw)。硬骨鱼类和其它脊椎动物的上、下颌分别被前颌骨(premaxilla)、上颌骨(maxil-la)和齿骨(dentary)等膜骨构成的次生颌(secondary jaw)所代替,而原来组成初生颌的骨块则退居口盖部或转化为听骨。第二对为舌弓(hyoid arch),包括背面一对舌颌骨(hyomandibular)、中部的一对角舌骨和位于腹面连接左、右角舌骨的单块基舌骨;舌颌骨的上端固着于脑颅,下端以韧带或通过其它骨块与下颌关连,鱼类以舌颌骨将下颌悬挂于脑颅的形式称为舌接式(hyostyl-ic)。第三至第七对是支持鳃的鳃弓(branchial arch)。硬骨鱼类的第五对鳃弓特化成一对下咽骨(hypopharyngeal bone),下咽骨上无鳃,其内侧在不同鱼类中长有数目、形状和排列方式各异的咽齿(pharyngeal teeth),常用于鲤科鱼类分类的依据。覆盖在鳃弓外侧并构成鳃腔的是3~4块鳃盖骨(opercular),其中以主鳃盖骨最大,它的上角前缘与舌弓背部的舌颌骨关接,可以在鱼类完成呼吸动作时,使主鳃盖骨的开关与口的闭启起到配合协调的作用。
2.脊柱(vertebral column)和肋骨(rib)脊柱紧接于脑颅之后,由一连串软骨或硬骨的椎骨关连而成,从头后至尾按节排列,取代了脊索的地位,成为对体轴强有力的支持及保护脊髓的结构。鱼类的椎骨完整,中央为椎体(centrum),椎体的两端凹入,是脊椎动物中最原始的双凹型(amphicoelous)椎体。相邻的2个椎骨之间彼此以前、后关节突(zygapophysis)关接,因而加强了椎骨的坚韧性和活动性。2个双凹型椎骨间所形成的球形腔内仍留有残存的脊索,并通过椎体正中的小孔道,使整条脊索串连成念珠状。脊柱的分化程度低,分为躯椎和尾椎两部分。每一躯椎由椎体、椎弓(又称髓弓neural arch)、髓棘(或称棘突neural spine)、椎体横突(parapophysis)等各部构成;尾椎则包括椎体、髓弓、髓棘、脉弓(haemal arch)和脉棘(haemal spine)等各部。两者在椎体上方的构造完全相同,但躯椎具有肋骨,而硬骨鱼类的肋骨还从两侧包围体腔起着保护内脏的作用;脉弓为尾动脉和尾静脉提供了通道。椎弓于椎体上方构成椎管,是容纳脊髓通过的管道。
(二)附肢骨骼(appendicular skeleton)鱼类的附肢骨骼包括鳍骨及悬挂鳍骨的带骨,而鳍骨又可分为奇鳍骨和偶鳍骨。
1.奇鳍骨 是一系列深埋于体肌肉的支鳍骨(担鳍骨pterygiophore),每个支鳍骨分为上、中、下3节,骨的上节支持着一根背鳍条或臀鳍条。尾鳍是鱼类游泳时的主要推进器官,最后几枚尾椎骨愈合成一根翘向后上方的尾杆骨,尾杆骨的上、下各有若干骨片或软骨片愈合而成的上叶和下叶,作为支持尾鳍鳍条的支鳍骨。
2.带骨和偶鳍骨(paired fin bone)悬挂胸鳍的带骨为肩带(pectoral girdle),由伸向背面的肩胛骨(scapule)、腹面的乌喙骨(coracoid)及匙骨(cleithrum)、上匙骨(supracleithrum)、后匙骨(postcleithrum)等组成,并通过上匙骨牢固地关连在头骨上。软骨鱼类的肩带位于咽颅的后方呈半环形,不与头骨或脊柱关连,只包括肩胛部和乌喙部两部分。肩带外侧有一与胸鳍连接的关节面,称肩臼(glenoid fossa)。绝大多数鱼类的胸鳍具有单列型(uniserial type)偶鳍骨(肺鱼类为双列型 biserial type偶鳍骨)。软骨鱼类的胸鳍内的支鳍骨有基鳍软骨(basal pterygium)和辐鳍软骨(radial pterygium),外侧为皮质鳍条,硬骨鱼类的支鳍骨趋于退化,常出现肩带直接关连鳞质鳍条的现象。
连接腹鳍的带骨为腰带(pelvic girdle),构造非常简单,位于泄殖孔前方,呈一字形的坐耻杆(ischiopubic bar),或由一对无名骨构成的三角形骨板。腰带两端通过关节面与腹鳍的鳍骨关连。雄性软骨鱼类的交配器叫作鳍脚(pterygopodium),是腹鳍内侧一块基鳍软骨特化所成的变形器官;硬骨鱼类腹鳍的支鳍骨与腰带愈合,或呈粒状介于无名骨和真皮鳍条之间。
三、肌肉系统
(一)头部肌肉 主要包括由脑神经控制活动的眼肌(extrinsic eyeball muscles)和鳃节肌(branchiomeric muscles)。咽区的8对肌节在未分化前清晰地呈按节排列,但由于头部骨骼发展,肌节退化消失或分化成结缔组织和舌肌等,只有最前面的3对耳前肌节保留下来,转变成眼肌。每个眼球上附有6条眼肌,其中上直肌、下直肌、内直肌和下斜肌由第一对肌节演变而来;上斜肌来自第二对肌节,外直肌由第三肌节变成。这些眼肌的收缩,可以使眼球往不同方向转动(图16-9)。眼肌是很稳定的肌肉,不仅鱼类如此,就是在脊椎动物各纲中也基本一致。

鳃节肌附生在颌弓、舌弓和鳃弓上,分别控制上、下颌的开关、鳃盖活动和呼吸动作等,受三叉神经(Ⅴ)、颜面神经(Ⅶ)、舌咽神经(Ⅸ)和迷走神经(Ⅹ)支配。上、下颌的出现引发了颌肌的发达。调节硬骨鱼两颌启闭的肌肉在头侧和腹面,司口关闭的是下颌收肌(m.adductor mandibularis)和咬肌(m.masseter)。位于头部腹面(左、右齿骨之间)的颏舌骨肌(m.geniohyideus)收缩时可使下颌降低,口即张开。
专司鳃盖提起的肌肉有鳃盖开肌、鳃盖提肌及舌颌提肌。鳃盖收肌和鳃条骨舌肌具有关闭鳃盖的作用,并使鳃条骨靠近身体,关闭鳃孔。
鳃肌常分化为背、腹两群,中间互不连接。背群主要有鳃弓提肌、鳃弓收肌、鳃间背斜肌,腹群有鳃间腹斜肌、鳃弓连肌、鳃弓牵引肌等。这些鳃肌的数目多、体积小、功能复杂而难于确定。
支配口、鳃盖开关和鳃弓活动的颉颃肌,有规律地交错收缩和宽息,便能形成水流不间断地进入口内和由鳃孔排出,完成摄食和呼吸动作(图16-10)。

(二)躯干肌 可分为大侧肌(m.lateralis)和棱肌(carinate muscle)二部分
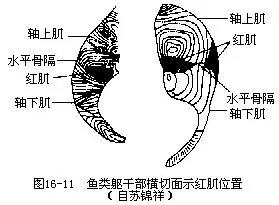
1.大侧肌 是鱼体上最大、最重要的肌肉,位于躯体两侧,由一系列被肌隔分开的锯齿形肌节组成,与圆口纲动物的肌节形式类同,是游泳前进的主要动力。每个肌节彼此以套叠形式而配置,因此在肌节弯曲处,形成具有若干同心圈的锥状构造,这在鱼体的横断面上显示得十分清晰。此外,沿水平体轴有一起于脊柱而外缘抵达皮下的水平骨隔(horizontal skeletogenous septum),将大侧肌分成上部的轴上肌(m.epaxialis)和下部的轴下肌(m.hypaxialis)。鱼类的轴上肌丰厚结实,是脊椎动物中最发达的类群,背鳍肌由此分化而来。轴下肌的肌层较薄,也无斜肌分化。多数鱼类的大侧肌可区分为性质和功能不同的2种肌肉,一种是条形的红肌或称浅层侧肌,由于肌肉内含有肌红蛋白(myoglobin)和大量血液而色泽暗红,含脂率高,位于水平骨隔上、下的躯干表层或脊柱两侧,能作长时间游泳的鱼类大多具有发达的红肌;另一种是白肌,是构成大侧肌的主要肌肉,不含肌红蛋白和脂肪,呈现白色,能行乏氧代谢,它是产生急速行动的物质基础,但缺乏持久的能力(图16-11)。
2.棱肌(carinate muscle)确切地说是一些支配升竖和降偃的纵形肌肉,如背鳍和臀鳍前、后的牵引肌及牵缩肌。此外,奇鳍在每一鳍条两侧基部各有3条束状肌肉;前侧部的竖肌、后侧部的降肌和介于两肌之间的倾肌,配合牵引肌和牵缩肌作用,控制奇鳍的升降。软骨鱼类无棱肌,故其奇鳍缺乏升降能力。
(三)附肢肌 胸鳍、腹鳍的浅层和深层均分布着由大侧肌分化而来的展肌(m.abductor)、伸肌(m.extensor)和收肌(m.adductor),支配偶鳍的向外伸展或内收。尾鳍肌比较复杂,每侧包括6块肌肉,控制尾的上曲下弯,并参与游泳时的推进运动。
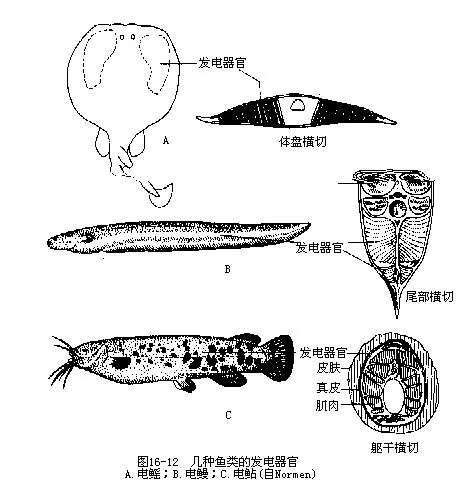
(四)发电器官 有些鱼类(鳐科、电鳐科、裸臀鱼科、电鳗科、瞻星鱼科Uranoscopoidae等)具有发电器官(electric organ),与御敌避害,攻击捕食、探向测位及求偶等活动有关。发电器官的位置因鱼种而不同(眼后、胸鳍基部、尾部)。其功能单位电细胞由肌细胞特化而成,电细胞集合成柱状作串联组合,发电器官的动作电位是由每个电细胞的电位差相加所得。每一电细胞的电位差约0.1Ⅴ。通常,产于南美洲河流中的电鳗尾部的发电器官约有电细胞6 000~8000个,放电时的总电压可高达600Ⅴ~800Ⅴ,电流由尾侧往前传向头侧。电鳐的发电器官位于胸鳍内侧,放电量仅100Ⅴ,电流的传导从背侧至腹侧。产于非洲河流中的电鲇(Malapterucuselectricus)放电量为400Ⅴ~450Ⅴ,但其发电器官并非肌肉变成,乃是真皮腺的特化(图16-12),裸臀鱼(Gymnarchus)和裸背鳗(Cymnotus)的放电量也极微弱。发电器官产生的电位取决于电细胞柱的数目,而电流强度则由所有电细胞柱横切面的总面积所决定。
四、消化系统
消化管包括口腔、咽、食管、胃、肠和肛门等。消化管由表及里由四层组成,即浆膜(serouslayer)、肌肉层(muscular layer)、粘膜下层(submucous layer)、粘膜层(mucous layer)。粘膜层的内壁上有纵行、网状等不同形式的褶皱或螺旋瓣(spiral valve),这是延缓食物在消化管的移行速度和增加吸收营养物质面积的适应结构。
鱼类的口腔和咽并无明显的界限,统称为口咽腔,内有齿、舌、鳃等器官,覆盖在口咽腔上的复层上皮富含单细胞粘液腺,但无消化腺及消化酶。许多鱼类不仅具有紧密贴合在上、下颌的颌齿(jaw teeth),也有附生于犁骨(vomer)上的犁齿(vomerine teeth)、腭骨上的腭齿(palatine te-eth)、舌骨上的舌齿(tongue teeth)和下咽骨上的咽齿(pharyngeal teeth)等。大多数硬骨鱼类的颌齿与咽齿的发展程度常成反相关的互补作用,即颌齿强大者,则咽齿不发达或退化,反之也然。鱼齿的形状各异,有犬齿形和圆锥形,也有臼齿形及门齿形,甚至还有前、后异形的颌齿,不同的齿型反映出鱼类在利用食料方面的差别。鲤科鱼类的口腔内缺乏颌齿。舌位于口咽腔底部,由基舌骨外覆有结缔组织和粘膜而成,只有舌端的游离部分具有肌肉,可作不同程度的上、下方向活动。有些鱼类的舌面布有味蕾,可能具有辨识食物的机能。鳃耙(gill rakers)是附生在鳃弓内侧两排并列的突起,是阻拦食物随水流出鳃裂的滤食结构;顶端尚有少量味蕾,也具有味觉器官的作用。鳃耙的数目、形状和疏密状况均与鱼的食性有关,如肉食性鱼类的鳃耙粗短而疏:鳡鱼13~15个,乌鳢10~13个,鳜鱼7~ 8个,青鱼18~20个;杂食或草食性的鲤鱼及草鱼其鳃耙数分别为14~18个及20~25个;吃浮游生物的鱼类,鳃耙细长而稠密,形成筛滤微小食物的网状结构,鳙鱼以浮游动物为食,第一鳃弓上就有680个左右鳃耙,而吃浮游植物的鲢鱼,鳃耙数可高达1700个,过滤面积约25cm2,使含有食物的水流在流入口咽腔时,通过鳃耙的筛滤作用,将食物截留,由粘液裹成食物团,利于吞嚥。食管(esophagus)短而环肌发达,壁厚,因布有味蕾,故对摄入的食物有选择及吐弃的功能。胃(stomach)是消化管最膨大的部分,前、后以贲门(cardiacportion)和幽门(pyloric portion)分别与食管及肠相通,有些鱼类(鲳鱼、鳜鱼)在幽门和十二指肠的交界处有许多突出的幽门垂(pyloric ceacum)。胃的机械性运动是蠕动,可使食物同胃液充分混合,混匀后的食物借胃壁的运动,通过幽门移入肠内。胃内壁具有胃腺(gastric gland),呈单盲囊状结构,分泌的胃液中包含盐酸、氯化物等无机盐和粘蛋白、消化酶等有机物,胃液分泌由食物直接刺激胃而引起。盐酸的作用在于激活胃蛋白酶原,供给胃蛋白酶所需要的酸性环境,使食物中的蛋白质变性,以利消化,同时还有一定的杀菌作用。
鱼类的食性复杂,胃液中所含的消化酶因鱼种而不同,也随其食性差异而不同。肉食性鱼类的胃蛋白酶活性甚高,主要是把蛋白质消化成蛋白和蛋白胨。非肉食性鱼类的胃蛋白酶少,但淀粉酶、糖原分解酶、麦芽糖酶的含量较多。捕食甲壳类和浮游动物的鱼类,胃内具有较强分解几丁质作用的几丁质分解酶。幽门垂开口于肠内,数量不一,玉筋鱼和鲻鱼最少,只有1~2个,鲷科和大多数蝶科鱼类为4个,红点鲑为20~22个,鳕鱼约380个,银鲳约600个,脂眼鲱(Etremeus micropus)约1000个,而斑(Clupanodon punctatus)的幽门垂甚至可达数千个之多。幽门垂内的粘膜具有众多褶皱和丰富的血管,并含有多种酶类,表明了这是一种既有辅助消化作用又兼有吸收功能的器官。
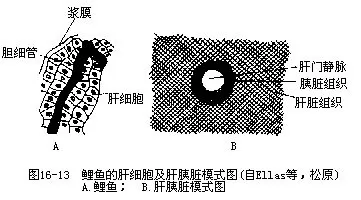
鱼类的肠管分化不明显,很难区分小肠和大肠,其长度随鱼种、食性和生长特性而不同。鲢鱼和草鱼等草食性鱼类的肠都很长,约为体长的6~7倍,有的鱼肠盘绕成圈几达体长的15倍,其中尤以吃硅藻的鲱鱼和鳀鱼最长。肉食性的乌鳢、鳜鱼、狗鱼、鳕鱼和鳀鱼的肠管最短,只及体长的1/3~3/4。有些鲤科鱼类的肠管长可随年龄增加而变长,例如,鲢鱼在体长5cm时,肠长是体长的3倍,当体长为6.7cm时,肠体之比增大成7.8倍;鳑鱼的生长与肠长同样也具有正相关的现象。鱼类的生长必然促进代谢量的相应增加,因此依靠增长肠管以扩展肠内的表面积。鱼类并无真正的肠腺,进行肠内消化的主要消化腺是肝脏和胰脏(pancreas)。肝脏的形状不一,一般分成2叶,但也有不分叶(鳗鲡)或呈3叶(鲣鱼、金枪鱼)、多叶(玉筋鱼)的。鲤科鱼类的肝脏呈弥散状分散在肠管之间的肠系膜上,因混杂有胰细胞而称肝胰脏(hepto-pancreas),但两种腺体的分泌物分别由胆管和胰管导入肠内(图16-13)。肝脏分泌的胆汁通过肝管贮存在胆囊中,再以胆管将胆汁输入肠内。胆汁能促进脂肪分解,使脂肪乳化,同时也有助于蛋白质的消化,促使某些蛋白质成分的沉淀。肝脏除制造胆汁外,还能把消化吸收的物质合成为糖原、脂肪和蛋白质。此外,对中间代谢、解毒作用、维生素及免疫物质的生成,都有重要作用。多数鱼类均有发达的胰脏,所分泌的胰液中含有胰蛋白酶、胰脂肪酶及淀粉酶。胰脏的消化酶需要
![]() 入肠内的盐酸而经常保持碱性状态,有利于胰液进行消化作用。有些鱼类的胰脏中埋有内分泌器官胰岛细胞,能分泌胰岛素(insu- lin),用作调节血醣的平衡。由此可见,肠是食物进行化学消化的主要场所,许多营养物质在这里被分解成简单成分。随着肠管的蠕动,把已消化了的食物吸入分布于肠壁上的血管和淋巴管中。未消化的食物残渣则经肛门排出体外。
入肠内的盐酸而经常保持碱性状态,有利于胰液进行消化作用。有些鱼类的胰脏中埋有内分泌器官胰岛细胞,能分泌胰岛素(insu- lin),用作调节血醣的平衡。由此可见,肠是食物进行化学消化的主要场所,许多营养物质在这里被分解成简单成分。随着肠管的蠕动,把已消化了的食物吸入分布于肠壁上的血管和淋巴管中。未消化的食物残渣则经肛门排出体外。
五、呼吸系统
鳃是鱼类的呼吸器官,位于口咽腔两侧对称排列。有些鱼类为适应某种特殊的生活条件,除鳃以外,还可通过皮肤(鳗鲡、鲇鱼、弹涂鱼等)、肠管(泥鳅)、鳃上器(攀鲈、斗鱼、乌鳢等)及气囊(肺鱼)等各种器官进行辅助呼吸,度过缺水乏氧的困难时期。此外,鳃还有排泄氮代谢废物及参与鱼体内、外环境的渗透调节等机能。
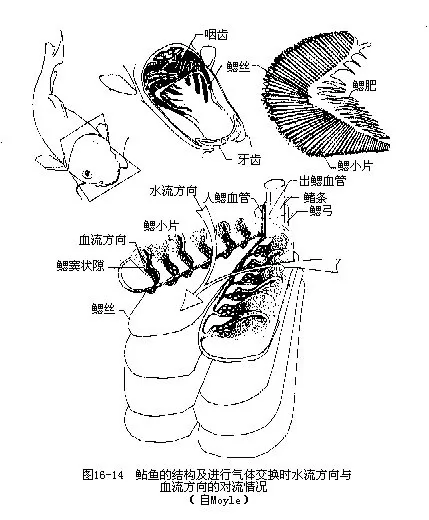
(一)鳃的构造 鱼类一般都具有5对鳃弓,前4对鳃弓的内缘着生鳃耙,最后一对特化成咽下骨,外凸面上长有2个并列的薄片状鳃片,每个鳃片叫做半鳃,长在同一鳃弓上的两个鳃片合称为全鳃,它们的基部彼此相连。软骨鱼类全鳃的2个鳃片之间有发达的鳃间隔(gill sep-tum),硬骨鱼类已退化消失。鳃弓之间形成5对鳃裂(gill cleft),鳃裂内、外分别开口于咽部及鳃腔(软骨鱼类直接开口体表),硬骨鱼类的鳃腔外覆有鳃盖骨(内缘附有鳃盖膜),以一总的鳃孔(gill opening)通向体外。鳃片由无数鳃丝(gill filament)排列构成,每一条鳃丝的两侧又生出许多突起为鳃小片(gill lamella),鳃小片由两层细胞组成,中间分布着丰富的微血管,是血液与外界水环境交换气体的场所。相邻鳃丝的鳃小片互相嵌合,成交错状排列,这种排列方式加上水流经鳃的方向与血流方向的对流配置(counter current principle),能保证血液与水流之间最大的气体交换量,从水中摄取的溶氧量可高达85%,若两者同向而流,则摄氧力只及对流状况的1/5(图16-14)。当鱼离开水域时,原来完全张开的鳃丝和鳃小片就彼此粘连,呼吸面积大大减小,无法获取充足的氧气,且鳃丝暴露在空气中,因水分蒸发而引起鳃片干燥,破坏了鳃的结构,使之失去呼吸机能而死亡。
(二)呼吸运动 鱼类在水中主要依靠口和鳃盖的运动完成呼吸动作。硬骨鱼类都有两对呼吸瓣,一对是附生于上、下颌内缘的口腔瓣(oral valve),闭嘴时可防止口中的水逆行流出,另一对附着在鳃盖骨后缘,即鳃膜,可阻止水从鳃孔倒流入鳃腔,同时也对口咽腔及鳃腔内的压力改变起着重要作用。
鱼类的呼吸运动是一个连续进行的过程,主要是运用下颌鳃部肌肉的收缩及口腔的协同动作,使口腔壁、鳃盖和鳃膜运动,改变口咽腔和鳃腔内部的压力,造成水流从口流入及由鳃孔流出,并在不断通过鳃的同时进行气体交换(图16-15)。一般认为,鱼类从口中进水的过程始于鳃膜紧闭的一瞬间。鱼口张开,鳃条骨展开而下沉,口咽腔的容积扩大,水被吸入口中,接着鱼口关闭,鳃盖骨往上提起,但此时鳃膜仍紧贴体表并盖着鳃孔,鳃腔的空间因此相应增大,内压减小,于是水流由口咽腔进入两侧的鳃腔,并通过鳃区。当水流通过鳃区进入鳃腔时,口腔瓣已经关闭,口咽腔在肌肉收缩的压力作用下,逐渐缩小,并往后波及到鳃盖部,造成鳃盖有力地拉下,使鳃腔内的压力增大,流入和积贮于鳃腔内的水便冲开紧贴在体表的鳃膜,从鳃孔流出体外。

然而,即使剪除硬骨鱼类的鳃膜,使之不能成为改变鳃腔内、外压力的调节结构,它们仍能从口中进水和自鳃盖后排水,从容地完成呼吸动作并在鳃区进行气体交换。不仅如此,有的金鱼品种(“翻鳃”)的鳃盖内缘并无鳃膜,人们可以在它提起鳃盖时看到鱼鳃,这就表明鳃膜在鱼类完成呼吸动作时,虽然具有一定作用,但是对于鱼类主动吞水在呼吸运动中的积极意义,同样不容忽视。金枪鱼类在持续地快速游泳时,口和鳃盖一直张开着,水流可以不停地进出于口和鳃孔,这一现象也能说明鳃膜在鱼类完成呼吸动作时,其作用是有局限性的。
延脑是鱼类呼吸中枢的所在地,在呼吸中枢控制下,通过第五、第九、第十对脑神经的支配,使颌部和鳃部的肌肉产生反射性的协调运动,完成呼吸动作。酸性水域会影响鱼从外界吸取氧气的能力,在碱性偏大的环境里同样不利于鱼类的呼吸。当pH值大于10或低于2.8时,均将造成鱼类呼吸器官的表面遭受损害。渔业生产上所说的“浮头”,就是由于池水中缺氧所引起,如不及时采取充气措施,常可造成大量池鱼窒息死亡,生产上叫做“泛塘”。在鱼类正常呼吸过程中,时常会出现呼吸节律被突如其来的短促的呼吸运动所打乱,这时有一部分水从口中吐出,同时有一部分水由鳃孔溢出,这种现象称为“洗涤运动”。洗涤运动的作用在于清除鳃上的外来污物,有利鱼类的气体交换。
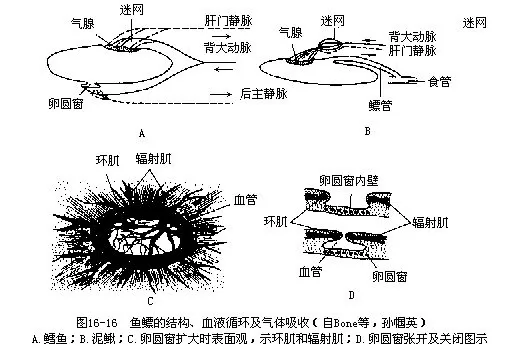
(三)鳔(gas bladder)绝大多数鱼类有鳔,少数种类(软骨鱼类、金枪鱼类和马鲛鱼等)无鳔,是次生现象。鱼鳔是位于肠管背面的囊状器官,鳔的内壁为粘膜层,中间是平滑肌层,外壁为纤维膜层。纤维膜层的细胞间因有小板状的鸟粪素结晶(guinine-crystals)而呈现白色。鳔因鱼种不同而分为1~3室,前、后室的间隔壁上有孔,便于鳔内气体流通和调节。根据鳔与食管之间是否存在相通的鳔管(pneumatic duct),可将鱼类分为两大类:一为有鳔管的管鳔类(physosto-mous),如鲤形目、鲱形目等,一为鳔管退化消失的闲鳔类(physoclistous),如鲈形目等,但是许多闭鳔类鱼类在刚孵化时是具有鳔管的,当通过鳔管将吞入的空气充满鱼鳔后即闭塞退化。鳔内的气体中除主含氮、氧和二氧化碳外,还有微量的氢、氩、氖、氦等。一般情况下,生活在浅水水域的鱼类,鳔内的含氧量甚低,以鲤鱼为例,氧含量仅占鳔内气体总量的2.42%,相当于它在4分钟内生活所需的氧气量,因而鱼鳔的呼吸机能是并无实际意义的。鱼鳔内的含氧量随同鱼的活动水层下降而逐渐升高,如鲂鱼在1m水深时,鳔的含氧量为16%,降至16m水深时,含氧量增高至50%,而在水深175m处活动的康吉鳗,鳔内的含氧量可高达87%。
鳔是鱼体比重的调节器官,它的机能是通过特有的气腺(gas gland)分泌气体以及卵圆窗(仅见于闭鳔类)或鳔管排放气体(管鳔类)而控制的。气腺位于鱼鳔前腹面的内壁上,因气腺的上皮细胞下方与稠密的微血管网相接而呈现红色,故又名红腺(red gland)。微血管网是一个由动脉微血管和静脉微血管组成的结构,称为迷网(rete mirabile),能往鳔内分泌气体。分布到气腺下的动脉微血管来自背大动脉或腹腔肠系膜动脉,静脉微血管离气腺后,经鳔静脉而注入肝门脉系统。由气腺分泌的气体进入鳔内是通过特定的生物化学反应实现的:静脉微血管内的血液携带着从气腺来的乳酸呼吸酶和碳酸酐酶,这些物质通过对流交换进入动脉微血管,乳酸在这里能促使溶解的气体释放出来,并穿过气腺的上皮细胞,进入鳔腔,而碳酸酐酶则可加速血液中碳酸的脱水作用,释出CO2进入鳔内,同时CO2的张力增加还能促进与血红蛋白结合的氧气分离开来,并穿透气腺上皮细胞渗入鳔中。气腺上皮细胞的这种穿透性是单向的,只允许气体向鳔内渗透而鳔中的气体则不能穿过气腺上皮细胞退回迷网。卵圆窗是闭鳔类硬骨鱼吸收鳔内气体的一种特有构造,位于鳔的后背方,窗壁很薄,厚度仅10μm~20μm,微血管丰富,以一小孔口与鳔体沟通,环肌和辐射肌围绕孔口,受迷走神经及交感神经支配,管理孔口启闭。在气体分泌和渗入鱼鳔的过程中,卵圆窗孔口的环肌一直处于收缩状态,孔口很小;当鳔内容积和压力增大需要吸收气体时,辐射肌就收缩,孔口扩大,可使鳔内的气弥散到卵圆窗周围的微血管中去。管鳔类则可直接通过鳔管从口或鳃孔排出鳔内的气体(图16-16)。
鳔内气体的分泌和吸收直接影响到鱼鳔的容积大小,在一定程度上可引起鱼体密度和比重的变化。不过,有鳔鱼类在水域中上升或下降活动都有一定范围,所能自由活动的水层是比较狭窄的,而且鳔内气体的分泌和吸收过程相当缓慢,不能快速地适应水压的变化,所以鳔的主要功能是使鱼体悬浮在限定的水层中,以减少鳍的运动而降低能量消耗。鱼类实现升降运动的主要器官则是鳍和大侧肌的运动。
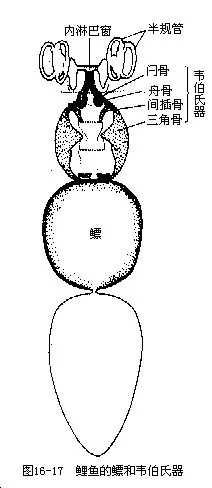
鱼鳔壁的四周分布着许多神经末梢,能感知声波及气体压力、水压的变化,并引起与此相适应的运动。鲤形目鱼类的鳔与内耳之间依靠由舟骨、间插骨、三脚骨等骨构成的韦伯氏器(webe-rian organ)联系,具有特殊的感觉功能(图16-17)。当外界声波传到鱼体时,鳔能加强这种声波的振辐,通过韦伯氏器可使鲤形目鱼类感受到高频率、低强度的声音,而鳔与内耳之间没有联系的鱼类,对声音频率的反应则不超过2000次/s~3 000次/s。
六、循环系统
循环系统包括液体和管道2部分。液体是指血液和淋巴液,管道为血管及淋巴管。鱼类的心脏构造和血液循环方式与圆口纲动物基本相同。
(一)心脏 位于鳃弓后下方的围心腔(pericardium cavity)内,后方以结缔组织的横隔与腹腔分开。由于心脏紧靠肩带,肩带从两侧和腹面包围心脏,使心脏得到很好的保护。心脏小,一般只占体重的1%左右(鲤鱼约0.11%,金枪鱼为0.3%,飞鱼为2.5%,软骨鱼类为0.6%~2.2%),由静脉窦、心房、心室等3部分构成。心室的前方有一稍为膨大的动脉圆锥(conus arte-riosus,是软骨鱼类心脏的一部分,能有节律地搏动)或动脉球(bulbus arteriosus,是硬骨鱼类腹大动脉基部扩大而成,不属于心脏的一部分,也无搏动能力)。静脉窦与心房之间有窦耳瓣,心房和心室之间有耳室瓣,心室与动脉球的交接处(即动脉圆锥所在地)有半月瓣(semilunar valve),所有这些瓣膜都具有提高血压和防止血液逆流的功能。
鱼类的心跳频率一般为每分钟18~20次。供给心脏营养的血液来自背大动脉或出鳃动脉以及锁骨下动脉的分支,离开心脏的血液注入前主静脉,再返回静脉窦。鱼体内的血量少,仅为体重的1.5%~3.0%,决不超过5%。
(二)血液循环 鱼类的血液循环方式属于单循环(single circulation),即从心脏压出的血液,经鳃区交换气体后,由出鳃动脉汇合成的背大动脉将多氧血运送至鱼体各部的器官组织中去,供给氧和各种必需的物质。离开器官组织的少氧血,又带着代谢废物或营养物质循着从小到大的静脉管道回流,最终汇流至心脏内,然后再开始新的一轮血液循环。单循环血液循环方式是与鱼类的心脏构造简单及用鳃呼吸密切相关的(图16-18)。

鱼的血液被心脏压出后,首先注入腹大动脉(aorta ventralis),再往前向两侧的鱼鳃发出4~5对入鳃动脉(arteria branchialis afferens)。入鳃动脉进一步分支成鳃丝动脉、鳃小片动脉并形成微血管网,交换气体就在鳃小片的微血管网进行。俟后,微血管依次汇合成出鳃小片动脉、出鳃丝动脉和出鳃动脉(a.branchialis efferens)。出鳃动脉中的血液汇入背大动脉,再由此发出许多动脉,将血液分流到鱼体各部。
大多数静脉都与动脉伴行分布。头部流回心脏的血液主要通过两侧的前主静脉(vena cardinalis anterior)及下颈静脉(vena jug-ularis inferior),分别接受来自脑、眼、鼻、颜面、鳃孔以及下颌和口咽腔的血液,向后与同侧的后主静脉(vena cardinalis posterior)共同组成一根粗大的总主静脉(vena cardinalis communis,即古维尔氏管Cuvier duct)开口于静脉窦。尾部及腹鳍的少氧血汇集到尾静脉(vena caudalis)而进入腹腔的肾脏中,分散为毛细血管而形成肾门静脉(vena portalis renalis),毛细血管与埋在肾脏中的后主静脉的毛细血管相连。后主静脉出肾脏后,接受来自各体节、背鳍、臀鳍、胸鳍的血液,并在心腹隔膜附近,与头部的前主静脉会合,通过总主静脉注入静脉窦。有些鱼类(丁及鳕鱼)的尾静脉可越过肾脏,直接与后主静脉相连而不形成肾门静脉。
鱼类的肝门静脉(vena portae hepaticus)很发达,凡是通过腹腔系膜动脉获得多氧血的消化器官,其回流的少氧血都注入肝门静脉,由此进入肝脏,分散成微血管,并与肝静脉(vena hepatica)的微血管相连。肝静脉微血管沿着血液流向由小而大,最后汇成2条粗大的肝静脉,注入静脉窦。从鳔和生殖腺来的少氧血也往往参与肝门静脉的组成。
(三)组织液和淋巴液 组织液来自毛细血管中的血液,是血管中的血液与血管外组织细胞之间物质交换的媒介。毛细血管壁具有通透性,它的末端血压高于外界液体的压力,所以血浆(滤去血细胞后的血液称为血浆)中的水及溶于水的小分子物质可通过毛细血管壁滤出,成为渗入组织细胞内的组织液。血浆中的一些大分子物质不易透过毛细管壁外,因此组织液中的蛋白质浓度低于血浆中的蛋白质浓度。蛋白质溶液产生胶体渗透压,而毛细血管中血浆的胶体渗透压高于组织液的胶体渗透压,这就是组织液在组织细胞内经过物质交换后重新被吸收进静脉毛细血管的动力。滤过和重吸收2种力的对比,是决定组织液生成或重新吸收的重要因素。
鱼类的淋巴系统不发达。身体各组织细胞间未被静脉毛细血管所吸收的少量组织液,可进入通透性极高而内压较低的淋巴管(lymphatic vessel),成为无色透明的淋巴液(lymph)。淋巴液来源于组织液,除不含红细胞和血液蛋白质外,其它成分与血液近似,它的流动方向为单向运行。淋巴管在最后一枚尾椎骨的下方,扩大成左、右相连的两个圆形淋巴心(Iymph heart),能不停地搏动,把淋巴液推向前行,最后流入后主静脉,参与血液循环。鱼类淋巴液的主要机能是协助静脉系统带走多余的细胞间液、清除代谢废料和促进受伤组织的再生等。
脾脏(spleen)脾脏是循环系统中的一个重要器官,是造血、过滤血液和破坏衰老红细胞的中心场所。脾脏位于胃的一侧或胃的后方,可分为外层红色的红髓和内层白色的白髓二部分。红髓制造红细胞及血小板,白髓生产淋巴球和白细胞。出入脾脏进行血液循环的血管分别为腹腔系膜动脉的分支脾动脉和脾静脉,最后血液注入肝静脉。脾脏内的微血管口径小,当衰老的红细胞通过管口时,易受损伤而导致死亡,其血红蛋白尤其是含铁部分则可被重新利用来制造新的红细胞。
七、神经系统和感觉器官
鱼类的神经系统由三部分组成,即中枢神经系统、外周神经系统和植物性神经系统。
(一)中枢神经系统(central nervous system)由脑和脊髓共同组成,并分别包藏在软骨或硬骨质的脑颅及椎骨的髓弓内。
1.脑 由端脑、间脑、中脑、小脑、延脑等五部分组成,结构比较简单,脑的体积也比其他脊椎动物小得多。据测定,鳗鲡的脑仅占体重的0.005%,而大多数鸟类和哺乳动物则为0.5%~2.0%。
端脑由嗅脑(rhinencephalon)及大脑(cerebrum)组成,嗅脑包括紧靠嗅囊的嗅球(olfactorybulb)和细长的嗅束(olfactory tract),往后与大脑相连。大脑中央有纵沟将其隔成左、右大脑半球(cerebral hemisphere),半球内各有一侧脑室。除软骨鱼类和肺鱼外,绝大多数鱼类的大脑背壁都很薄,无神经组织,主要由嗅神经组成特有的古脑皮(paleopallium),即嗅脑。腹面有纹状体(striatum corpora),是许多神经细胞集中形成的脑组织所在处。大脑是鱼类的嗅觉和运动调节中枢,损伤或切除其大脑后,除失去嗅觉外,还会降低游泳能力及繁殖行为的调节作用。间脑位于大脑后方,内部有第三脑室,背面中央突出一条细长的脑上腺,常被中脑的一对视叶所遮盖。间脑腹面的前方有视神经,并形成神经交叉(optic chiasma),交叉后有一椭圆形的漏斗(infundib-ular)及与其相连的脑垂体(hypophysis)。漏斗两侧有一对下叶,它的后方是血管囊(vascularsac),这个结构在深海鱼类尤为发达,有人认为它是一种压力感受器。间脑第三脑室的上皮中具有光敏感细胞,可因鱼体感受光度增加引起皮肤内黑色素细胞扩散,使体色变黑;间脑的一些神经元有分泌机能,这种现象称为“神经分泌”,其分泌物不但具有温度调节、心血管调节和摄食调节效应,而且还有控制脑垂体分泌的作用,间接地影响着鱼类的繁殖活动。中脑是位于间脑上方的一对椭圆形球体,又名视叶,是所有脊椎动物的视觉中心;脑内有中脑腔,为连接第三、第四脑室的通道。视觉冲动从视网膜通过视神经传导到中脑,终止于中脑背侧部,若切除中脑,则鱼类顿失视觉。此外,中脑与侧线、体表、小脑、延脑等也有神经联系,这些联系对调整鱼体位置及控制运动的机能有关。小脑位于中脑后方,是身体活动的主要协调中枢,具有维持鱼体平衡、掌握活动的协调和节制肌肉张力等作用;小脑两侧有耳状或球形的小脑鬈,与内耳及侧线器官的关系密切,所以鱼类的小脑又是听觉和侧线感觉的共同中枢。延脑位于脑的最后部,是多种生理机能和感觉的中枢。延脑的前部有面叶及位于其两侧的迷走叶,是感受口内和皮肤表面味觉的中枢。延脑体前宽后窄,背面覆有脉络丛(choroid plexus),膜下为三角形的第四脑室,脑室后方与脊髓内的神经管腔相通。延脑是鱼类的听觉、皮肤感觉、侧线感觉和呼吸的中枢,同时它还是调节色素细胞作用的中枢,能使身体的色素细胞收缩,引起皮肤颜色变淡(图16-19)。
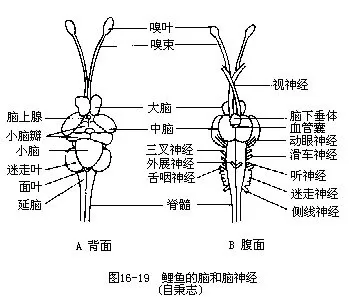
2.脊髓 是一条扁圆形的柱状管,包藏于椎骨的髓弓内,前面与延脑连接,往后延伸到最后一枚椎骨。脊髓由前向后逐渐变细,但因出现胸鳍和腹鳍而在其相应部位略显膨大。背、腹面分别具有背中沟和腹中沟,以此将脊髓分成左、右两半。围绕在脊髓神经管腔四周成蝶形的灰质(gray matter),是神经原本体,灰质周围为白质(white matter),里面只有神经纤维。
脊髓是中枢神经系统的低级部位,以脊神经与机体的各部相连系。白质是上行和下行神经束的通道,传导感觉和运动的神经冲动,把鱼体组织器官与脑的活动互相连结起来。灰质部分有神经元,可完成最基本的反射活动,所以脊髓是鱼体和内脏反射的初级中枢所在处,但它的活动都是在中枢神经系统的高级中枢部位支配下进行的。
(二)外周神经系统(peripheral nervous system)由中枢神经系统发出的脑神经和脊神经组成,其作用是通过外周神经将皮肤、肌肉、内脏器官所来的感觉冲动传递到中枢神经,或由中枢向这些部位传导运动冲动。
1.脑神经(cranial nerves)鱼类有脑神经10对,其名称及分布部位在无羊膜类各纲动物中大致相同。
(1)嗅神经(olfactory nerve):神经元的细胞体分布在嗅囊的粘膜上,由细胞体轴突集合成的嗅神经终止端脑的嗅叶或大脑。嗅神经的功能专司嗅觉。
(2)视神经(optic nerve):神经元的细胞体位于眼球的视网膜上,由轴突合成的视神经穿过眼球壁和眼窝,在间脑腹面形成视神经交叉,入间脑而最后抵达中脑。视神经专司视觉。
(3)动眼神经(oculomotor nerve):由中脑腹面发出,分布到眼球的上直肌、下直肌、内直肌和下斜肌,与滑车神经和外展神经共同支配眼球的活动。
(4)滑车神经(trochlear nerve):由中脑侧背面发出,穿过眼窝壁,分布到眼球的上斜肌上。这是唯一由中枢神经系统背面发出的一对运动神经。
(5)三叉神经(trigeminal nerve):发自延脑的前侧面,在通出脑颅前神经略为膨大,称为半月神经节(semilunal ganglion)。三叉神经在神经节后分成4支:深眼支、浅眼支、上颌支和下颌支。深眼支分布到鼻部粘膜和吻部皮下。浅眼支与面神经的浅眼支在基部并合,一起分布到头顶及吻端的皮肤上。上颌支沿口角分布到上颌。下颌支由口角分布至下颌。三叉神经既支配着颌的动作,也接受来自吻部、唇部、鼻部及颌部的感觉刺激。
(6)外展神经(abducens nerve):由延脑腹面发出,穿过眼窝壁分布于眼球的外直肌上。
(7)面神经(facial nerve):从延脑侧面发出,与三叉神经的基部接近,关系也甚密切。面神经分为3支:浅眼支、口支和舌颌支。浅眼支与三叉神经的浅眼支合并,一起分布到吻部。口支穿过脑颅,分布到上颌和口腔顶部。舌颌支最粗大,分支可达舌弓、舌颌骨、下颌骨、鳃盖骨及鳃条骨等。面神经支配头部和舌弓的肌肉运动,并接受来自皮肤、触须、舌部和咽鳃等处的感觉刺激。
(8)听神经(auditory nerve):由延脑侧面发出,与三叉神经、面神经、舌咽神经的基部彼此靠拢,分布至内耳的半规管、椭圆囊、球状囊及壶腹上,感知听觉和平衡感觉。
(9)舌咽神经(glossopharyngeal nerve):从延脑侧面发出而紧挨听神经之后,穿过前耳骨到达第一鳃弓。它的基部有一神经节,节后分为鳃裂前支和鳃裂后支,分布到口盖部、咽部、鳃裂的壁上及头部侧线系统。
(10)迷走神经 (vagus nerve):发自延脑侧面最粗大的一对脑神经,由此分出3支:鳃支、内脏支及侧线支,分布到第一至第四鳃弓、心脏、消化器官、鳔及侧线系统上。迷走神经的机能是支配咽喉部和内脏器官的活动,并感受咽部的味觉、躯干部的皮肤感觉及侧线感觉等(图16-20)。
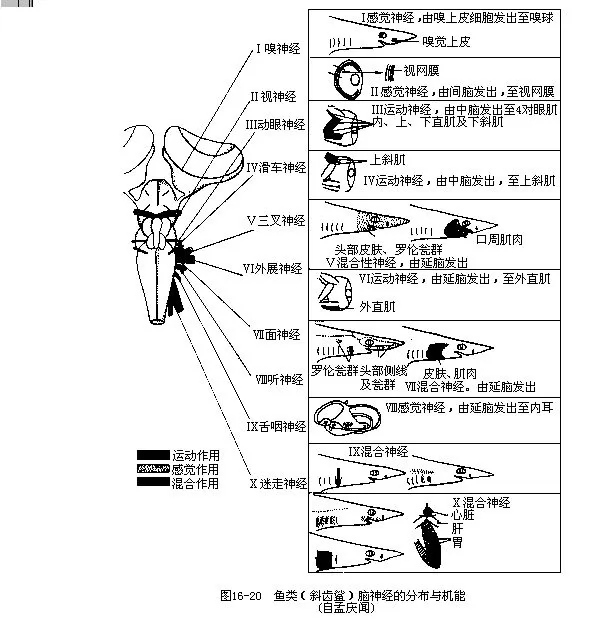
鱼类的10对脑神经中,嗅神经、视神经及听神经为纯感觉神经,仅由导入的感觉神经纤维构成,分别与嗅、视、听觉发生联系;动眼神经、滑车神经和外展神经是纯运动神经,只包含运动神经纤维,用于支配动眼肌的活动;三叉神经、面神经、舌咽神经及迷走神经均为混合神经,兼有感觉和运动两种神经纤维,主要与咽弓、内脏的感觉和运动有关。
2.脊神经 脊髓具有明显的分节现象,每节发出一对左、右对称的脊神经与外周相联系。每一脊神经包括一个背根(dorsal root)和一个腹根(ventral root)。背根连于脊髓的背面,腹根发自脊髓的腹面。背根内主要包括感觉神经纤维,腹根主含运动神经纤维。背根有感觉作用,它的神经纤维来自皮肤和内脏,负责传导周围部分的刺激至中枢神经系统,靠近脊髓处各有一膨大的脊神经节(spinal ganglion),联系周围部分及脊髓;腹根的神经纤维分布到肌肉和腺体上,用以传导自中枢神经系统发出的冲动到周围各反应器。
脊神经的背根和腹根在穿出椎骨的椎间孔之前互相并合,并在椎间孔外分成3支:第一支是背支(dorsal ramus),分布到鱼体背部的肌肉及皮肤;第二支是腹支(ventral ramus),分布到腹部的肌肉和皮肤上,这两支都兼含感觉神经纤维及运动神经纤维,各自分管鱼体背、腹部肌肉的运动和感觉;第三支是内脏支(visceral ramus),分布到胃、肠和血管等内脏器官上,同样含有2种神经纤维,支配其感觉和运动(图16-24)。脊神经在肌节上的分布有彼此重叠现象,即每一肌节可同时接受2~3个腹根支配,而每个腹根又可支配几个肌节的作用。因此如果只有一小段脊髓连同其腹根受损时,能引起几个肌节的肌力衰退,但还不至于造成一个肌节完全瘫痪。部分脊神经的腹支在偶鳍的相应部位形成神经丛(nerve plexus),这是脊椎动物在进化过程中出现成对肢体后发展而成的形态特征,用以加强胸、腹鳍的活动支配能力。

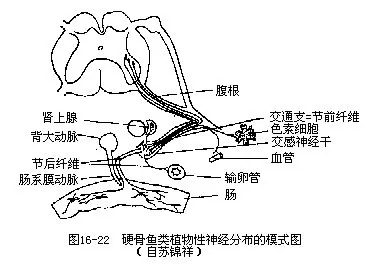
(三)植物性神经系统(vegetative nervous system)是专门支配和调节内脏平滑肌、心脏肌、内分泌腺、血管扩张和收缩等活动的神经,与内脏器官的生理活动、新陈代谢有着密切的关系。植物性神经也由中央神经系统的脑或脊髓发出,但并不直接到达所支配的器官,而是先通过具白色髓鞘(mylin sheath)的节前纤维(preganglionic fibres,构成白交通支white connecting strand),到达一个交感神经节(sympathetic ganglion),然后再由灰暗色无髓鞘的节后纤维(postganglionic fi-bres)分布到各器官(图16-22)。
植物性神经系统可分为交感神经系统(sympathetic nervous system)和副交感神经系统(para-sympathetic nervous system),其神经纤维同时分布到各种内脏器官,产生颉颃作用,器官在两种对立作用的制约下,才能维持其平衡和正常的生理功能。鱼类的植物性神经系统尚处于初级阶段,还不及高等脊椎动物那样发达和完善。大多数都有沿脊柱两侧按节排列的交感神经节,以及由此以神经纤维前、后相连而成的交感神经干(sympathetic trunk)。交感神经干前伸至延脑两侧,与三叉神经、面神经、舌咽神经、迷走神经等紧密相连。头部副交感神经包括在发自动眼神经和由延脑发出的迷走神经中,循着这2对神经分布到眼球虹膜、消化管、心脏和鳔等器官上。
(四)感觉器官(sensory organ) 鱼类的感觉器官主要包括皮肤感受器、侧线感觉器、听觉器官、视觉器官、嗅觉器官和味觉器官等
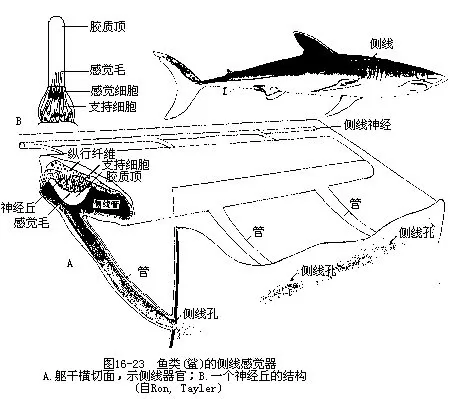
1.侧线系统(lateral line system)是鱼类特有的皮肤感觉器官,呈管状或沟状,埋于头骨内和体侧的皮肤下面,侧线管以一系列侧线孔穿过头骨及鳞片,连接成与外界相通的侧线。侧线管内充满粘液,感觉器就浸埋在粘液里。感觉器一般由一群感觉细胞和一些支持细胞组成,称为神经丘,感觉细胞具有感觉毛和分泌机能,其分泌物在整个感觉器的外部凝结成胶质顶,感觉神经末梢分布于感觉细胞之间。当水流轻击鱼体时,水压通过侧线孔,影响到管内的粘液,并使感觉器内的感觉毛摆动,从而刺激感觉细胞兴奋,再通过神经将外来刺激传导到神经中枢(图16-23)。支配头部侧线的是面神经和舌咽神经,而支配躯干部侧线的是迷走神经的侧线支。侧线能感受低频率的振动,具有控制趋流性的定向作用,同时还能协助视觉测定远处物体的位置,故在鱼类生活中具有重要的生物学意义。
软骨鱼类除侧线外,吻部还有特殊的皮肤感受器—罗伦氏壶腹(ampulla of Lorenzini),由罗伦瓮、罗伦管和管孔三部分组成。罗伦瓮是一个基部膨大的囊状结构,内有腺细管和感觉细胞,面神经的神经末梢分布其间;罗伦管是由罗伦瓮通出的管道,末端以管孔开口于皮肤表面(图16-24)。罗伦氏壶腹是水流、水压、水温的感受器,也是电感受器官,能检测出低限至0.01μm/cm的电压。
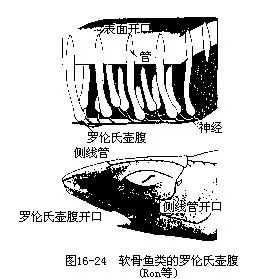
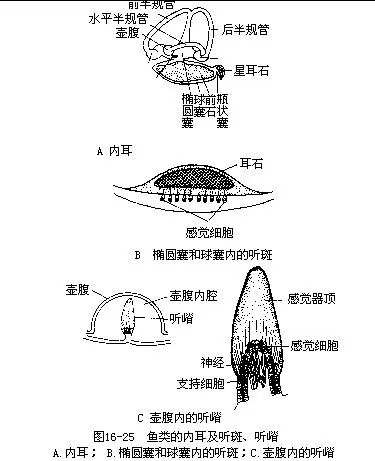
2.听平衡觉器官(auditory organ)鱼类的听平衡觉器官是一对内耳,因其结构复杂而称膜迷路(membrane labryinth),包藏于脑颅听囊内的外淋巴液中,膜迷路里充满着内淋巴液。每侧的内耳都包括上、下二部分:上部是椭圆囊(utriculus)及与其连通的3个半规管,管的一端膨大成壶腹(ampulla);下部是球囊(sacculus),球囊后方有一突出的瓶状囊(lagena),这些囊内有石灰质的耳石(otolith)3~5块,其中以球囊中的翦石(sagitta)体积最大。椭圆囊、球囊和壶腹内的感觉上皮,分别形成听斑(acoustic spot)和听嵴(acoustic crista),与听神经的末梢相联系,是鱼类平衡和听觉器官中的主要感受部位(图16-25)。当鱼体移位时,耳石对听斑和听嵴的压力起了变化,内淋巴液的压力也随之发生改变,于是感觉的信息通过听神经传递到中枢神经系统,引起肌肉反射性运动。膜迷路上部的椭圆囊和半规管是鱼体平衡机制的中心,而球囊和瓶状囊内的听斑能感受声波,并通过听神经将外界的声浪传到脑,产生听觉。
前半规管
(自苏锦祥)
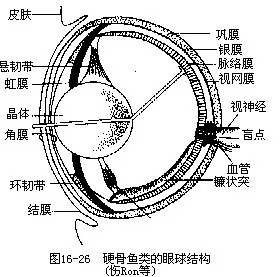
3.视觉器官(optic organ)多数鱼类缺乏活动性眼睑,有些鲨鱼(真鲨、双髻鲨、猫鲨、皱唇鲨等)的下眼睑内有瞬膜或瞬褶(nictitating fold),可向背上方移动,遮蔽鱼眼。鱼的眼球呈球状,具3层被膜:外层是软骨质或纤维质的巩膜(sclera),巩膜在前方形成透明而扁平的角膜(cor-nea),有保护眼及避免因磨擦而遭受损伤的作用。中层是脉络膜层(choroid),由自外向内的银膜(agentea)、血管膜(vascular coat)和色素膜(uvea)组成。银膜为鱼类所特有,呈银色而含鸟粪素,可将射入眼球的微弱光线反射到视网膜上;血管膜与色素膜互相紧贴难以分辨。脉络膜往前延伸成虹膜(iris),虹膜中央的孔即瞳孔(pupil)。眼球的最内层为视网膜(retina),是产生视觉作用的部位,由数层神经细胞组成,内含司光觉的圆柱细胞(rod cell)和感知色觉的圆锥细胞(conecell);视神经分布到视网膜上,视神经通出处无视觉作用,称盲点(blind point)。
眼球内有透明细胞构成的晶体(crystalline lens),角膜与晶体之间,以及晶体与视网膜之间分别有水样液(aqueous humor)和玻璃液(vetreous humor),前者有反光能力,而玻璃液则能固定视网膜的位置,使透过它的光线落到视网膜上。晶体大而圆,无弹性,背面藉悬韧带连接在虹膜上,紧挨于角膜后方,使鱼眼只能看到较近处的物体。镰状突(falciforme process)是硬骨鱼类调节视距的特有结构,为一膜质的垂直隆起,一端附着盲点,另一端以韧带与附着在晶体腹面的晶体缩肌(又称铃状体 Haller’s campanula)相连,通过该肌的伸缩能稍稍移动晶体的位置,调整视距,适应观察较远处的物体,但最远的视距一般不超过15m(图16-26)。
鱼眼一般位于头部两侧,紧靠吻部前方是不能见物的无视区。无视区之前则为两眼都能见物的双眼视区,也是鱼类视觉最清晰和对物体距离具有精确感觉的区域。一侧鱼眼能见物体的范围,称为单眼视区(图16-27)。

八、排泄系统
鱼类通过肠管和鳃排出未能消化的食物残渣及一部分代谢的废物,但大部分代谢废物都是以尿的形式由肾脏滤出,并通过输尿管排出体外。排泄系统由一对肾脏及输尿管组成,其功能除排泄尿液外,在维持鱼体内体液的适当浓度、进行渗透压调节方面也具有重要作用,以保证机体适应所处的环境。
(一)肾脏及泌尿机能肾脏紧贴于腹腔背壁,是一对坚实而呈块状的泌尿器官。有些硬骨鱼类的肾脏前端尚有不具有泌尿机能的头肾(head kidney)。肾脏由许多肾小体(renal corpuscle,或称马氏体 Malpighian body)构成,肾小体包括肾小球(glomerulus)和肾小管(renal tuble)两部分。肾小球是背大动脉分支在肾小管的肾口旁形成的一个毛细血管团;肾小管的前端凹入,由两层扁平上皮细胞构成杯状的肾小球囊(renal capsule,或称鲍曼氏囊 Bowmen’s capsule),将肾小球包入其内。肾小球囊的囊壁分内、外两层,2层之间有一狭小腔隙,称为肾囊腔,与肾小管的管腔相通。半渗透性的肾小球囊从毛细血管的血液内滤泌的尿液,经肾小管后段的吸水作用,曲折盘行汇集到总的输尿管。尿在肾脏中的生成过程是连续不断的,生成后经输尿管流入膀胱(urinarybladder)暂时贮存起来,积聚到一定量时,再经泌尿孔(urinary pore)一次性排出体外。
淡水鱼类肾脏内的肾小球数量明显多于海洋鱼类,这种特点与它们具有高渗性的体液有关。淡水鱼类在进行鳃呼吸及取食的同时,也摄入了过量水分,为维持正常范围的体液浓度,就必须通过众多肾小球的滤泌作用,增大泌尿量而排除体内的多余水分。
鱼类尿液中的含水量达95%以上,此外还含有少量肌酸、肌酐、尿酸等难于扩散的氮分解物,以及钠、钾、钙、镁、氯、硫酸盐和磷酸盐等无机盐。
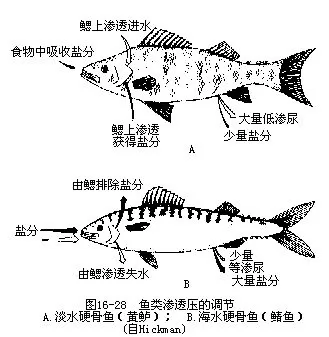
(二)渗透压的调节淡水和海水的含盐度相差极大,但是分别栖息于2种不同水域中的鱼类,其体液所含盐分浓度却并无显著差异,这就表明鱼类具有调节渗透压的机能。淡水鱼类体液的盐分浓度一般高于外界环境,为一高渗溶液,以血液冰点下降(△℃)表示其渗透压,约为-0.57,而淡水则接近于0℃(海水为-2.0℃)。按渗透原理,体外的淡水将不断地通过半渗性的鳃和口腔粘膜等渗入体内,但肾脏可借助众多肾小球的泌尿作用,及时排出浓度极低几乎等于清水的大量尿液,保持体内水分恒定。淡水鱼类在尿液的滤泌和排泄过程中,丧失的盐分很少,这是因为肾小管具有重吸收作用,将滤泌尿液中的盐分重新吸收回血液内。此外,有些鱼类还能通过食物或依靠鳃上特化的吸盐细胞从外界吸收盐分,这对鱼类维持渗透压的平衡,也具有重要的作用。如把淡水鱼置于海水中,则会造成组织失水而体内积贮过量盐分、血液粘滞性提高、血细胞沉降速度减慢,最后导致死亡。
海洋鱼类体液内的盐分浓度比海水略低,为一低渗性溶液。按渗透原理,体内水分将不断地从鳃和体表向外渗出,若不加以调节,可因大量失水而死亡。为维持体内、外的水分平衡,鱼类除了从食物内获取水分外,尚须吞饮海水,然而吞饮海水的结果又造成了盐分浓度在鱼体内的增高。为减少盐分的积聚,海鱼把吞下的海水先由肠壁连盐带水一并渗入血液中,再由鳃上的排盐细胞将多余的盐分排出而把水分截留下来,使体液维持正常的低浓度。海洋鱼类肾脏内的肾小体数量比淡水鱼类少得多,甚至完全消失,以此达到节缩泌尿量和水分消耗的目的(图16-28)。软骨鱼类用另一种方式调节渗透压以适应海水生活,它们的血液中因含有2%左右的尿素而浓度高于海水,不致产生失水过多现象。尿素是软骨鱼类在海水中使之保持体内水和盐分动态平衡的主要因子,当血液内尿素含量偏高时,从鳃区进入的水分就多。进水量增多后稀释了血液的浓度,排尿量随之相应增加,因而尿素流失也多。当血液内尿素含量降低到一定程度时,进水就会自动减少,排尿量相应递减,于是尿素含量又开始逐渐升高。
综上所述,淡水鱼类和海洋鱼类由于生活在环境条件不同的水域中,所以二者分别通过其独特的途径进行渗透压调节。因而海洋鱼类都不能进入淡水生活,反之亦然。
九、生殖系统
由性腺(精巢和卵巢)及输送生殖细胞的生殖导管组成。进行体内受精的鱼类,其雄性还有特殊的交配器。性腺左右成对而对称,但黄鳝和银汉鱼(Atherina hepsetus)等只有左侧的发达。性腺由系膜悬系于腹腔背壁。
(一)两性异形 鱼类一般都是雌雄异体(gonochorism),在鲱鱼、鳕鱼、黄鲷、狭鳕和几种鱼中发现有雌雄同体(hermaphroditism)现象。两性鱼甚至还有自体受精能力。此外,黄鳝、剑尾鱼、鱼等少数种类尚有性逆转现象,即性腺的发育从胚胎期一直到性成熟期都是卵巢,只产生卵子,经第一次繁殖后,卵巢内部发生了改变,逐渐转变成精巢而呈现出雄鱼特征。
一般在外形可用于区分性别的只有软骨鱼类腹鳍内侧的鳍脚、食蚊鱼臀鳍鳍骨特化而成的交配器( clasper)和雌性鳑鱼类由生殖孔伸出体外的产卵管等(图16-29)。而表现于两性异形的第二性征则是多方面的:既有雌鱼体大于异性10~30倍以上的角鳑和康吉鳗,也有雄鱼体略大于雌性的黄颡鱼和棒花鱼等;雄性鱼(Zacco)和马口鱼(Opsaurichthys bidens)的前部臀鳍条显著延长;雄银鱼的臀鳍上方有一横列大鳞;雄泥鳅的胸鳍约与头长相等,而雌性则甚短小;雄鱼的腹鳍后缘抵达肛门,雌鱼则不然;雌、雄鳜鱼的生殖孔分别为横形和圆形。
很多鱼类在进入生殖期时,雄鱼常出现某些与繁殖活动有关的第二性征,俟生殖期结束后即消失或复原,其中较明显和引人注目的是婚色(nuptial color)、珠星(nuptial organ)等。雄性棒花鱼在生殖期间,全身变黑,背鳍也变得比平时更为宽大;雄性泥鳅于后背部加厚,俯视时略呈方形体;雄性鳑鱼类于臀鳍下缘出现艳丽的红、黄、黑三色镶边;雄性中华多刺鱼的腹棘内侧出现小锯齿,并在半透明的腹棘内呈现鲜蓝色;雄性斗鱼在体侧出现绚丽而呈蓝宝石形的小圆斑,这些婚色的出现都是由于生殖腺分泌的性激素在血液中作用的结果。珠星是雄鱼表皮细胞特别肥厚和角质化的产物,外观为白色坚硬的锥状体,主要分布在吻、颊、鳃盖及胸鳍上,而香鱼和雅鲁鱼的追星几可遍及全身。
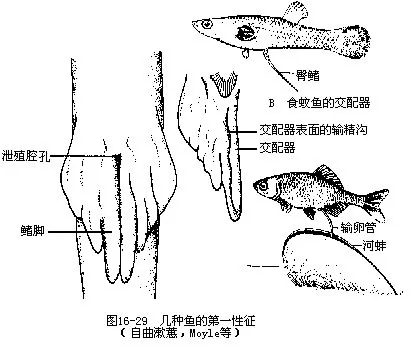
(二)生殖器官
1.雄性 精巢在性成熟时呈乳白色,表面匀净细腻,俗称鱼白,而在一般情况下为淡红色。精巢圆柱形或成盘曲的细带状,在生殖期达到最大体积,通常位于腹腔左、右两侧,彼此分开,但有时也可在后部互相接触或合而为一。精巢表面有腹膜所形成的外膜,内由不规则排列的壶腹形腺体或辐射状排列的叶状腺体构成,这些腺体就是精子的生长发育和成熟处。精子可从精巢的背面或底部的输出管注入输精管。
输精管是脊椎动物中自鱼类才开始出现的生殖导管。硬骨鱼类的输精管由精巢外膜往后延续而成,与肾脏无任何联系,这种现象在脊椎动物中是绝无仅有的。左、右输精管常在后段连合,以生殖孔开口于肛孔和泌尿孔之间,精子排至水中营体外受精。雄性软骨鱼类的输尿管兼有输精管的作用,管的后部膨大成贮精囊(seminal vesicle),往后分别导入尿殖窦(urogenital sinus),经尿殖乳头末端的孔开口于泄殖腔,精子由泄殖腔再通过鳍脚上的沟进入雌鱼生殖导管内。营体内受精。
2.雌性 鱼类的卵巢有两种类型:游离卵巢和封闭卵巢。游离卵巢的表面裸露,不包有腹膜形成的卵囊膜,卵巢里面有许多滤泡,每个滤泡内有1枚卵子,滤泡在卵子成熟时能分泌溶解酶,溶解滤泡,卵子即突破而出排入腹腔,借体壁肌收缩,经输卵管腹腔口进入输卵管,软骨鱼类和肺鱼类的卵巢均属于这一类型。硬骨鱼类的卵巢为封闭卵巢,卵巢外被腹膜形成的卵囊膜包围,并往后延伸变窄成输卵管(oviduct),成熟的卵不排入腹腔,而是落进卵巢的卵巢腔内,由此直接输送进输卵管。未发育成熟的卵巢常呈透明的条状,一旦成熟即变为长囊形而几乎充塞整个腹腔,颜色则由白转为黄色,但有些鱼类(红鳍鲌、鲇鱼)也可呈现绿色或橘红色(大麻哈鱼)等。
(三)性腺发育及卵的结构 掌握鱼类性腺各发育阶段的形态特征是鉴别鱼体是否达到性成熟的指标,因而对养殖业和进行人工繁殖工作具有十分重要的实践意义。鱼类性腺的发育一般分为6期(图16-30)。
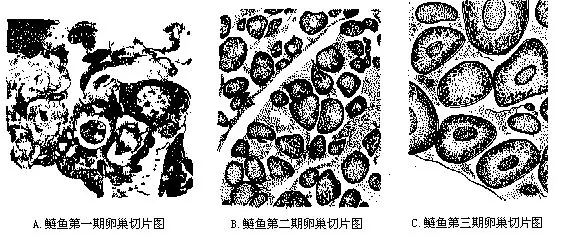
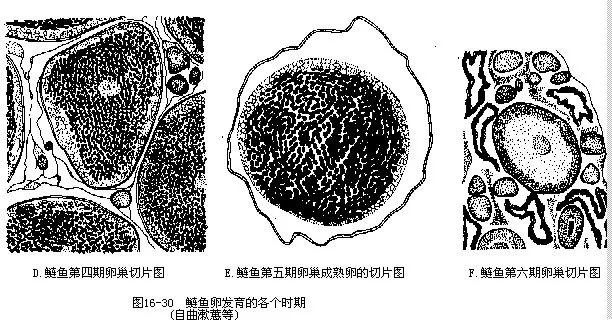
Ⅰ期性腺为一线状体,紧贴于腹腔背壁,肉眼无法分辨其雌雄。
Ⅱ期卵巢呈扁带状,表面有血管分布而呈浅粉红色或淡黄色,放大镜下已可看清卵粒,卵径90μm~300μm。精巢仍为细带状,血管不明显,故大多呈浅灰色。
Ⅲ期卵巢青灰色或黄白色,肉眼能辨认其卵粒,但卵粒互不分离,卵径250μm~500μm。精巢因血管发达而呈粉红色或淡黄色。
Ⅳ期血管十分发达,整个卵巢显示浅黄色或粉红色,卵形饱满,但在卵巢内因彼此挤压而略呈不规则状,卵径800μm~1500μm。精巢转为乳白色,表面血管清晰。
Ⅴ期用手触摸腹部有松软感,提起雌鱼或轻压鱼腹时,成熟的卵子从生殖孔中可自然流出。以同样方式处理雄鱼,也有大量粘稠的乳白色精液溢出体外。Ⅵ期卵巢因产卵后体积大大缩小,显得松软而空虚,由于血管充塞,故外观呈紫红色。精巢萎缩成细带形,浅红色或淡黄色。
有些鱼类在人工饲养条件下,由于缺乏或不能满足其某些必要的生殖条件,即使亲鱼已经生长到性成熟阶段,却不能进行繁殖活动。这时,饲养人员可对亲鱼注射脑下垂体液、绒毛膜促性腺激素,促进其产卵和排精活动。
除软骨鱼类和食蚊鱼(柳条鱼)、海鲫、汤氏鲪、绵鳚等营卵胎生(ovoviviparity)或胎生(vivi-parity)繁殖外,绝大多数鱼类的繁殖方式为卵生。鱼类产圆形的端黄卵(telolecithalegg),卵黄含量丰富,细胞质不多,分布在卵子周围和核的附近,受精后集中到动物极形成胚盘。各种鱼类的卵径大小不一,虎鱼的卵径仅0.3mm~0.5mm,矛尾鱼为85mm~90mm(重320g,是硬骨鱼类中最大的卵)而鼠鲨的卵径为150mm~220mm。鱼卵依其本身的比重不同,可分为浮性卵和沉性卵两大类,前者具有油球,色泽透明,产出后即漂浮于水面。淡水鱼类大多产沉性卵,卵内不含油球,产出后常沉入水底。
鱼卵表面为一薄层的卵膜,外部包有角质或胶质构成的壳膜。在动物极附近的壳膜上有一小孔,称卵膜孔,似可释放出一种由糖蛋白或粘多糖组成的受精素(fertilizin),以诱导精子游趋此孔而达到受精目的。
第三节 鱼纲分类
现存鱼类约22000种,分为软骨鱼类(Chondrichthyes)和硬骨鱼类(Osteichthyes)两大类群。
一、软骨鱼类
内骨骼全为软骨的海生鱼类;体被盾鳞;鼻孔腹位;鳃间隔发达,鳃孔5~7对。鳍的末端附生皮质鳍条。歪型尾。无鳔和“肺”。肠内具螺旋瓣。生殖腺与生殖导管不直接相连;雄鱼有鳍脚,营体内受精。全世界约800种,我国产190多种,绝大多数分布于热带及亚热带海洋,包括两个亚纲。
(一)板鳃亚纲(Elasmobranchii) 体呈梭形或盘形。鳃孔5~7对,各自开口于体外而无鳃盖;上颌不与颅骨愈合。雄性仅有位于腹鳍内侧的鳍脚。共2总目。
1.鲨形总目(Selachomorpha) 体呈梭形,鳃孔侧位,故又称侧孔总目(Pleurotremata)。胸鳍与头侧不愈合;背鳍背位;歪型尾(图16-31)。
(1)六鳃鲨目( Hexanchiformes):我国产六鳃鲨科( Hexanchidae)的3属3种,最常见的是分布于黄海、渤海的扁头哈那鲨(Notorhynchus platycephalus),体长4m~5m,重200kg~300kg。鳃孔7个。底栖生活,游泳缓慢,吞食小鱼和甲壳动物。卵胎生。肝含油量达65%~70%,可制鱼肝油。东海和南海偶而也见灰六鳃鲨(Hexanchus griseus)。
(2)虎鲨目(Heterodontiformes):头大吻钝,眼上棱起显著,有鼻口沟;前方的牙尖细,后方的牙平扁呈臼齿状。背鳍2个,前方各具一枚鳍棘;有臀鳍。全世界只有虎鲨科(Heterodontidae)1属8种,我国产2种,即体纹宽而纹间距较大的宽纹虎鲨( Herterodontus japonicus)和体纹窄而纹间距小的狭纹虎鲨(H.zebra),分别分布于黄渤海及南海。


(3)鲭鲨目(Isuriformes):背鳍2个,无硬棘;有臀鳍。全世界有4科7属14种。较有代表性的种类是体长约15m的姥鲨(Cetorhinus maximus)、尾鳍长度超过体长1/3的长尾鲨(Alopiasvalpinus)和俗称大白鲨的噬人鲨(Carcharoldon carcharias)等。
(4)须鲨目(Orectolobiformes):有鼻口沟或鼻孔开口至口内。前鼻瓣常有一鼻须或喉部具一对皮须。最后2~4对鳃孔位于胸鳍基底上方。第一背鳍与腹鳍相对或位于其后。全世界有3科12属约29种,我国产3科8属12种,常见种类有头侧有皮瓣的日本须鲨(Orectolobus ja-ponicus)、豹纹鲨(Stegostoma fasiatum)和世界上最大的鲸鲨等。
(5)真鲨目(Carcharhiniformes):眼有瞬膜,是世界上软骨鱼类中属、种最多的一个类群,共8科47属200余种,我国产4科23属60多种,常见种类有体具斑点的梅花鲨(Halaelurusburgeri)、星鲨(Mlustelus)、颌齿斜行排列的斜齿鲨(Scolidon sorrakowah)和锤头双髻鲨等。
(6)角鲨目(Squaliformes):背鳍2个,大多有硬棘。无瞬膜;有喷水孔;鳃孔位于胸鳍基底前方;缺乏臀鳍。我国产角鲨科(Squalidae)1属6种,沿海常见的有短吻角鲨(Squalus breviros-tris),吻短圆钝,鼻孔接近吻端;背部棕褐色,无斑。
(7)锯鲨目(Pristiophoriformes):头平扁,吻长似剑状突出;鼻孔前方有一对皮须;具瞬膜及喷水孔。无臀鳍。本目只有锯鲨科(Pristiophoridae),我国产日本锯鲨(Pristiophorus japonicus),体长可达4m,底栖生活,食甲壳动物和蠕虫等,分布于黄海、东海和南海。
(8)扁鲨目(Squatiniformes):是鲨形总目中唯一体型平扁的类群。胸、腹鳍扩大,且彼此接近;背鳍2个,形小而位于尾部上方。本目仅扁鲨科(Squatinidae)1属,我国产2种,常见种为日本扁鲨(Squatina japonica),俗称琵琶鲨。
2.鳐形总目(Batomorpha)体形扁平,鳃孔腹位,又名下孔总目(Hypotremata);胸鳍前部与头侧相连;背鳍常位于尾上;无臀鳍;尾鳍或有或无。全世界有4目20科约430种,我国产约80种(图16-32)。
(1)锯鳐目(Pristiformes):吻狭长而平扁,似剑状突出,边缘具尖利的吻齿。本目只有锯鳐科(Pristidae),分布于热带和亚热带沿岸海区,有些种类也可进入江河生活。我国南海和东海产尖齿锯鳐(Pritis cuspidatus),鱼体最长可达9m,常用剑状的吻锯击毙或刺伤追食对象。
(2)鳐形目(Rejiformes):吻圆钝或突出,侧缘无吻齿。本目主要包括体盘呈犁状的犁头鳐(Rhinobatus hynnicephalus)、体盘呈团扇形的团扇鳐(Platyrhinus sinensis)和体似斜方形的鳐鱼(Raja)等。
(3)鲼形目(Myliobatiformes):胸鳍往前延伸到达吻端,或前部分化为吻鳍或头鳍。腹鳍前部不分化成足趾状构造。我国产约31种,主要种类有尾长加鞭而无背鳍的类(Dasyatis)、体盘宽阔而尾甚短小的燕(Gymnura japonica)、胸鳍与吻鳍在头侧相连的鸢鲼(Maliobatis tobi-jei)和蝠鲼(Mobula japonica)等。类中的赤不仅是北方的常见种,而且也生活在珠江水系直至广西龙州一带。
(4)电鳐目(Torpediniformes):体盘椭圆形,皮肤光滑柔软;头侧与胸鳍之间的皮下具有特化而成的发电器官。我国有2科4属8种。常见种类有产于南海的黑斑双鳍电鳐(Narcine macu- late)和日本单鳍电鳐(Narke japonica)等。
(二)全头亚纲(Holocephali) 体表光滑或偶有盾鳞。鳃腔外被一膜质鳃盖,后缘具一总鳃孔。背鳍2个,第一背鳍前有一强大硬棘,能自由竖立或垂倒。雄性除腹鳍内侧的鳍脚外,尚有腹前鳍脚及额鳍脚。全世界有1目3科近30种。我国产5种,最常见的为银鲛科(Chimaeridae)的黑线银鲛(Chimaera phantasma),俗名海兔子,吻圆锥形,体银灰色,头和背侧部暗褐色,侧线下方有一黑色纵带,以无脊椎动物及小鱼为食。
二、硬骨鱼类
骨骼大多由硬骨组成;体被骨鳞或硬鳞,一部分鱼类的鳞片有次生性退化现象;鼻孔位于吻的背面;鳃间隔退化,鳃腔外有骨质鳃盖骨,头的后缘每侧有一外鳃孔。鳍的末端附生骨质鳍条,大多为正型尾。通常有鳔,肠内大多无螺旋瓣;生殖腺外膜延伸成生殖导管,二者直接相连。无泄殖腔和鳍脚,营体外受精。硬骨鱼类包括2亚纲。
(一)内鼻孔亚纲(Choanichthyes)本亚纲鱼类的口腔内具有内鼻孔(choana);有原鳍型的偶鳍,即偶鳍有发达的肉质基部,鳍内有分节的基鳍骨支持,外被鳞片,呈肉叶状或鞭状,故又称肉鳍亚纲(Sarcopterygii)。肠内有螺旋瓣。共有2总目。

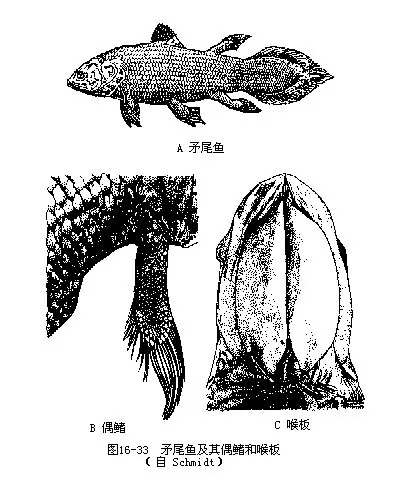
1.总鳍总目(Crossopterygiomorpha)这是一类出现于泥盆纪的古鱼,也是当时数量最大的硬骨鱼类。具有中轴骨骼是一条纵行的脊索而不存在椎体、颏下有一块喉板(gular plate)、肠内有螺旋瓣等一系列原始特征。早期的总鳍鱼类都栖居于淡水中,有鳃、鳔和内鼻孔,能在气候干燥和水域中周期性缺氧期间用鳔呼吸空气。同时凭借肌肉发达的肉叶状偶鳍支撑鱼体爬行。长期以来,总鳍鱼类一直被认为已于中生代末期的白垩纪时完全绝灭,但是1938年12月22日却在非洲东南沿海哈隆河河口水深70m首次捕获一尾体长1.8m、重95kg的总鳍鱼,并依据其尾形定命为矛尾鱼(Latimeria chalumnae),该标本被保存于东伦敦博物馆内(图16-33)。以后又在科摩罗伊斯兰联邦共和国科摩罗群岛附近的海域中陆续捕得150~200尾矛尾鱼。这是总鳍鱼类唯一现存的残留种,隶属于腔棘鱼目(Coelacanthiformes)、矛尾鱼科(Latimeriidae)。这些古鱼的孑遗已成为动物界最珍贵的活化石之一。
矛尾鱼全长1m~2m,重13kg~80kg。体被平列的圆鳞而带金属蓝色;肉叶状偶鳍的表面覆有鳞片,主要鳍骨与陆生脊椎动物的肢骨极为相似;尾鳍呈特殊的三叶式矛头形。生活在水深50m~550m的海洋中,游泳迅捷;齿呈颗粒状,在口缘形成齿板,肉食性;卵胎生。由于矛尾鱼类在适应海洋生活的进化过程中,摒弃了它们远祖用鳔呼吸的习性,所以鳔已变为充有结缔组织和脂肪的两个囊状结构,内鼻孔也随之发生次生性外移而在口腔中消失。
2.肺鱼总目(Dipneustomorpha)本总目是与古总鳍鱼类亲缘关系较近的同时代的鱼类,两者的主要区别是肺鱼类口腔中有内鼻孔、偶鳍内具双列式排列(辐鳍骨列于基鳍骨的两侧)的鳍骨和高度特化而适于压碎软体动物硬壳的迷齿。肺鱼类一方面具有某些原始特性:上、下颌由腭骨、翼骨、犁骨、夹板骨、隅骨等构成而无次生颌;脊索终生保留,组成椎骨的骨片直接连在脊索上,椎体尚未形成;心脏前有动脉圆锥;肠内具螺旋瓣;尾鳍为原型尾(protocercal tail)。另一方面还具有与其生活环境相适应的特化性状或进化特征。例如,肺鱼类的鳔不论在发生、构造或是呼吸机能上,都与陆生脊椎动物的肺十分相似。鳔有鳔管与食道相通,外界的空气可通过口腔和内鼻孔直接进入鳔内。与鳔呼吸相联系的心脏及血液循环方式也发生了相应变化:心房被不完全的房间隔分成左、右两半,在鳔内经气体交换后的多氧血流回心房的左半边,从身体回归的少氧血流进心房的右半边,是脊椎动物进化过程中首次出现包括体循环和肺循环的双循环。本总目在世界各地曾有过广泛的分布,但现生种类仅2目3科5种,并被隔离分布于南美洲、亚洲和大洋洲,我国四川省境内也出土过肺鱼化石。
(1)单鳔肺鱼目(Ceratodiformes):体形侧扁,胸鳍粗阔;体鳞大。鳔不成对。生活在不干涸的水域中,也不进入蛰眠状态,常躺卧于水底,捕食甲壳动物、软体动物和蠕虫等。9、10月间繁殖,产卵于水生植物中间,幼鱼无外鳃,发育毋需经过变态。现存澳洲肺鱼科(Ceratodidae)的澳洲肺鱼( Neoceratodus forsteri),分布于澳大利亚昆士兰。
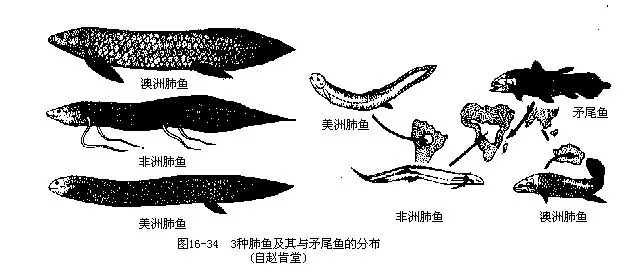
(2)双鳔肺鱼目(Lepiosireniformes):体呈鳗形,胸鳍鞭状或较狭短;体鳞小,埋于皮下。鳔成对。旱季或枯水期即钻入水底淤泥中,用皮肤分泌的粘液,在鱼体周围作茧而进入夏眠,此时仅用鳔进行呼吸。雨季来临时,水位升高,肺鱼即可苏醒破茧而出活动,并进行鳃呼吸。以水底无脊椎动物为食,也吃一部分水生植物。卵产于水底的孔穴中,幼鱼有外鳃,发育需经变态。本目包括2科:美洲肺鱼科(Lepidosirenidae),偶鳍细小而短,奇鳍低矮;幼鱼的外鳃存在期短,仅1属1种,即美洲肺鱼(Lepidosiren paradoxa),产于南美洲亚马逊河流域;非洲肺鱼科(Protopteri-dae),偶鳍细长似鞭,奇鳍高;外鳃保留于整个幼鱼期,本科共1属3种,较常见的是非洲肺鱼(Protopterus annectens),分布于非洲中部(图16-34)。
(二)辐鳍亚纲(Actinopterygii)本亚纲鱼类的各鳍均由真皮性的辐射状鳍条支持。体被硬鳞、圆鳞或栉鳞,或裸露无鳞。无内鼻孔。种类极多,占现生鱼类总数的90%以上,共包括9总目、36目。产于我国的有8总目、26目。
1.硬鳞总目(Ganoidomorpha)本总目是鱼类中古老类群的残余种,除了具有硬骨鱼类的主要特征外,仍留有一些原始性状:体被外覆硬鳞质(ganoid)的菱形硬鳞;心脏具动脉圆锥;肠内有螺旋瓣;尾鳍为歪型尾;颏部常有喉板。包括4目,即鲟形目、多鳍鱼目、弓鳍鱼目和雀鳝目等(图16-35)。(1)鲟形目(Acipenseriformes):体形似鲨,吻长,口腹位,有喷水孔。躯干部有5行纵列的骨板,或皮肤裸露而仅在歪型尾的上叶列有少数硬鳞性质的叉状鳞。内骨骼为软骨,仅于头部具有膜质硬骨;髓弓、脉弓、间髓弓、间脉弓等骨片并不愈合,脊索发达,无椎体。我国产2科。
鲟科(Acipenseridae)体被5行纵列骨板,口具伸缩性,前方有4条吻须。幼鱼期有齿,成长后则消失无迹。本科是北半球的淡水鱼或溯河性鱼类,鳇鱼(Huso dauricus)的最大体长超过5m,重1000余kg,口裂大,几达头侧,鳃盖膜与峡部不连;5~7月产卵,食鱼和无脊椎动物。鲟属(Acipenser)鱼类的口小,鳃盖膜与峡部相连,代表种类有新疆额尔齐斯河及伊犁河产小体鲟(A.ruthenus)、东北产施氏鲟(A.schrenckii)、长江中上游产长江鲟(A.dabryanus)。中华鲟(A.sinensis)为江海洄游性鱼类,成鱼每年5~6月溯河到长江上游产卵,孵化的稚鱼在江内栖居一段时期后又返回海里。1983年,长江水产研究所在葛洲坝获得中华鲟催青育苗成功,并开始人工放流,为扩大淡水养殖业增加了新的珍贵鱼类品种。
白鲟科(Polyodontidae)体表裸出,无成行骨板,仅尾鳍上叶有叉状鳞。口内有细齿。本科仅2属2种,呈不连续分布:一种是产于长江和钱塘江的白鲟(Psephurus gladius),白鲟全身灰白色,别名象鱼,头吻均长,往前延伸呈剑状突出,有一对短须,是我国特有的大型珍稀鱼类;另一种分布于北美密西西比河的匙吻白鲟(Polyodon spathula)。
(2)多鳍鱼目(Polypteriformes):背鳍分离为一列小鳍,故得名多鳍鱼,每个小鳍前方各有一枚鳍棘。胸鳍有肉质的基叶,然其鳍骨却与总鳍鱼明显不同。尾鳍为圆形的正型尾。鳔分2叶,内多分隔,开口于食道腹面。幼鱼期有外鳃。主要种类为具有腹鳍的多鳍鱼(Polypterus bichir)和体形似鳗而腹鳍退化的芦鳗(Calamoichthys congicus)等。多鳍鱼类全产在非洲热带河流中,栖于水下的泥底,性情凶猛,以小鱼、甲壳动物、昆虫为食;当河湖接近干涸时,可用鳔直接呼吸空气。
(3)弓鳍鱼目(Amiiformes):体被圆形硬鳞,但不含闪光质,内骨骼大多为硬骨。颏下有一大形喉板。鳔内分成许多小室,为辅助呼吸器官。中生代中期曾是弓鳍鱼类全盛时期,而现存种类仅一种,即弓鳍鱼(Amia clava)。弓鳍鱼两颌具细齿,肠螺旋瓣和动脉圆锥极度退化,并出现了动脉球,这些特点表明该鱼虽然还具有一些原始构造,但其演化地位已经比较接近真骨鱼类。生活于北美的静水和缓流中,食物以鱼为主。雄鱼在繁殖季节常选择水草丛生的湖边浅滩构筑粗糙的圆形鱼巢,供雌鱼产卵。此外,它还有卫护受精卵和稚鱼的习性。
(4)雀鳝目(Lepidosteiformes):体被不含闪光质的菱形硬鳞。无喉板及喷水孔。鳔有呼吸作用。现生种类仅雀鳝科(Lepidosteidae),代表动物为扁口雀鳝鱼(Lepidosteus platystomus),体大,长达2m左右,两颌前伸成长吻,齿尖利,是北美和中美洲淡水湖中的肉食性鱼类。
2.鲱形总目(Clupeomorphe)腹鳍腹位,鳍条一般不少于6枚;胸鳍基部位置低,接近腹缘;鳍无棘。圆鳞。主要有以下诸目(图16-36)。
(1)海鲢目(Elopiformes):是硬骨鱼类中的低等类群,有些还保留动脉圆锥和喉板等原始特征。稚鱼为“柳叶体”型,在个体发育中有变态。我国产3种,主要分布于南海和东海。常见种类有北梭鱼(Albula vulpes)和大海鲢(Megalops cyprinoids)等。
(2)鲱形目(Clupeiformes):鳍无棘,背鳍单个。体被圆鳞,无侧线。本目中有许多种类是很有经济价值的海鱼,在世界渔业中占有重要的地位。代表种类有:


鲱科(Clupeidae)体侧扁,鳞片容易脱落。口较小,口裂不超过眼的后缘;齿细弱;腹缘通常有锯齿状棱鳞。大多是集群性的中上层鱼类,其中包括世界渔业中最著名的鲱鱼类和沙丁鱼类,它们的产量几乎占到世界渔产量的22%。我国的重要种类有鲱鱼(Clupea pallasi)是黄海的重要经济鱼类。鲥鱼(Maccrura reevesi),4~6月由海中溯河洄游至珠江、闽江、钱塘江和长江产卵。繁殖后亲鱼回归海洋,幼鱼则留在江河中肥育到当年秋季才降河洄游入海,是我国的名贵优质鱼类。鳓鱼(Ilisha elongata),味鲜美,由此加工而成的广东曹白鱼、浙江糟鲞和虾子鲞鱼均为颇具盛名的佳品。
鲑科(Engraulidae)上颌骨后延超过眼的后缘;吻突出,覆于口上;臀鳍大多较长。广泛分布于热带和温带水域中,体形不大,但数量甚多,是沿海常见的经济鱼类,其中尤以凤鲚(Coiliamystax)、刀鲚(C.ectenes)和鲑鱼(Engraulis japonicus)最为重要。凤鲚的臀鳍与尾鳍连接,故又称凤尾鱼,胸鳍上方有游离鳍条6根,为河口性鱼类,春季鱼群进入咸淡水区域产卵,亲鱼生殖完毕后仍回海中生活,常在长江下游形成较大渔汛。刀鲚的生活习性与凤鲚相似,但是在太湖还有陆封型的小型刀鲚,也是淡水捕捞的对象。鲑鱼的臀鳍短,不与尾鳍相连;腹部无棱鳞,是小型的中上层海鱼,常集成大群作昼夜垂直移动,为我国北方和东南沿海的经济鱼类,加工后的幼鱼干即“海蜒”,常作为煮汤食用。
(3)鲑形目(Salmoniformes):体形和特征与鲱形目相似,但背鳍后方常具一脂鳍(adiposfin);有侧线。大多栖居于北极和高纬度水域内,我国黑龙江、内蒙古、新疆和西藏境内均有分布。本目包括在世界渔业中占重要地位的鳟鲑鱼类,主要的有:
鲑科(Salmonidae)口裂大,齿锥形;有脂鳍。侧线完全。为北半球定居性和进行溯河洄游的冷水性鱼类。大麻哈鱼(Oncorhynchus keta)是我国鲑科鱼类中的主要捕捞对象,生殖鱼群每年秋季由太平洋上溯至黑龙江、乌苏里江、松花江产卵,并于此期间停止摄食,体色由银色转变成暗灰色,雄鱼身上出现红棕色斑点,两颌变为钩状。亲鱼生殖后不久死去,受精卵于翌年孵化,仔鱼生长到体长50毫米时开始降河入海,在海里生活3~5年,俟性成熟时成群洄游至江中繁殖。分布在东北的还有乌苏大麻哈鱼(O.masou)和驼背大麻哈鱼(O.gorbuscha)等。哲罗鱼(Huchotaimen)的臀鳍条7~10枚,是栖于黑龙江、乌苏里江、松花江及新疆额尔齐斯河的冷水性大型鱼类,最高体重可达50kg以上,性凶猛,以鱼类及水生昆虫为食。繁殖期间体色变红,溯河上游到水质清沏的砾质河床掘穴产卵,新疆地区俗称大红鱼。
胡瓜鱼科(Osmeridae)体鳞小,侧线不完全,是分布于我国东北的小型鱼类,常见的是池沼公鱼( Hypomesus olidus),肉多骨软,味腴美,新鲜时有清香浓郁的黄瓜味,营养价值高。1983年起,辽宁省已将此鱼移殖于宽甸县的各水库中,作为组织对外出口的生产基地。
银鱼科(Salangidae)生活于淡水、咸淡水或海水中的小型鱼类,细长而透明,除雄鱼臀鳍上方有一行鳞片外,周身裸露无鳞。头平扁,吻部尖突;口裂大,两颌、口盖及舌上均有细齿。鱼卵表面有卵丝。我国产8属13种,统称面条鱼或面杖鱼,产量较大的有太湖银鱼(Neosalanxtaihuensis)和大银鱼(Protosalanx hyalocranius),前者于1981年移殖至云南滇池,而大银鱼也在1988—1992年先后于内蒙古、北京地区移养成功,并取得一定生产规模的经济效益。
狗鱼科(Esocidae)吻突出似鸭嘴,上颌骨无齿。背鳍位置与臀鳍相对,无脂鳍,是分布在北半球的冷水性淡水鱼类。我国产东北狗鱼(Esox reicherti),背部青铜色,缀有许多大形黑斑,栖息于黑龙江水系及内蒙古达赍湖,性凶贪食,年产量大,有经济价值;白斑狗鱼(Esoxlucius)的体侧有淡蓝色斑,仅产于新疆额尔齐斯河流域。
(4)灯笼鱼目(Myctophiformes):口大,两颌、腭骨及舌上均有能倒伏的尖齿。具脂鳍。鳍无棘。我国有11科,以狗母鱼科(Synodidae)中的蛇鲻(Synodus elongata)和龙头鱼(Harpodon ne-hereus)产值最大。龙头鱼分布于南海、东海和黄海,肉质松软,加工成的干制品称为龙头烤,是浙江沿海人民嗜食的水产品。
3.鳗鲡总目(Anguillomorpha)体呈鳗形。腹鳍腹位或缺失;背鳍及臀鳍的基底长,与尾鳍相连。个体发育有变态。是栖息于热带和亚热带水域中的海洋鱼类,只有少数种类(鳗鲡An-guilla和头齿鳗Pisodonophis)可进入淡水中。成鱼在沿岸浅海内生活,肉食性,生殖时到深海产卵。仔鳗变态后游向近岸。共3目,我国产鳗鲡目(Anguilliformes)9科,主要有(图16-37):
鳗鲡科(Anguillidae)体呈蛇形,鳞小,埋于皮下成席纹状排列。共1属2种,常见种鳗鲡(Anguilla japonica)是典型的降河性洄游鱼类,它们在江河中生长到六龄时,开始入海繁殖,排卵完毕和卵受精后亲鱼全部死亡。仔鱼无色透明,形似柳叶,称叶状仔鳗(Leptocephalus),具有针状细齿,捕捉微小生物为饵,游泳能力弱,常在随海水漂流到近岸期间变态为线状幼鳗(或称玻璃鳗 glass eel),此时细齿已消失,代之以圆锥形的新齿,鳗龄约3岁,准备开始上溯进入江河。离水时可用皮肤进行辅助呼吸。
海鳗科(Muraenesocidae)口大,吻长,舌附于口底,颌齿中杂有大形犬牙。体无鳞,侧线完全。本科中的海鳗(Muraenesox cinereus)是重要的经济鱼类之一,性甚凶猛,游泳迅速,白天隐伏于岩穴和海底泥沙中,夜间外出捕食,有季节性洄游。海鳗鳔也为名贵食品。

4.鲤形总目(Cyprinomorpha)是比较低等的硬骨鱼类。腹鳍腹位,有些种类(鲇形目)有脂鳍;鳔有管与食道相通;具韦伯氏器。广布于世界各洲,大部分种类生活于淡水,但以热带和亚热带水域中最多,可生活在高山、平原、江河、山溪、激流中。包括许多重要的经济鱼类和养殖鱼种,全世界约有5000种,分鲤形目和鲇形目2个目。
(1)鲤形目(Cypriniformes):体被圆鳞或裸露。许多种类口内无齿,但下咽骨有发达的咽齿。全世界有3000余种,分布几乎遍及各地。我国产6科,重要的有(图16-38):
胭脂鱼科(Catostomidae)口小,下位;咽齿一行,数目多而排列成栉状。唇厚,密生乳突。北美产的种类多,我国仅有分布在长江和闽江流域的胭脂鱼(Myxocyprinus asiaticus),背鳍高大而长,鳍条50~57枚,栖于水的中下层,以无脊椎动物为食。1975年,浙江淡水研究所与四川宜宾鱼种场合作开展了胭脂鱼的人工繁殖及移养,获得成功。
鲤科(Cyprinidae)是鱼类中种类最多的一个科,约有2000余种。主要特征是上颌口缘由前颌组成,咽齿1~3行。鳔的前部大多没有膜囊或骨囊包围。无脂鳍。本科鱼类是我国淡水天然捕捞以及池塘和大水面养殖的主要对象,有经济价值的不下400种,产量约占全国渔产总量的1/3。重要的鲤科种类有:草鱼(Ctenopharyngodon idellus),吻较宽钝,无口须;咽齿2行,栉状;体棕黄色,鳞缘黑色;草食、生长迅速和鱼肉质佳,历来是我国的淡水养殖鱼种。青鱼(Mylo-pharyngodon piceus)外形很像草鱼,但吻较尖,咽齿一行,臼齿状;除腹部外,身体各部和鱼鳍均呈青黑色;生活于水的中、下层,以螺、蚌和蚬等为食;主要分布在长江以南的平原地区。鲢(Hypophthalmichthys molitrix),头长约占全长的1/4;眼下侧位;鳃耙细密,呈海绵状;体银白色,故称白鲢;中、上层鱼类,活泼善跳,主要食物是浮游生物和各种碎屑。

鳙(Hypophthalmichthysnobilis),外形酷似鲢鱼,但因背侧的体色较暗,并有不规则黑点而俗称花鲢;头长约为全长的1/3,故又名胖头鱼;栖于水的中、上层,主要分布在长江和珠江流域,与草鱼、青鱼、鲢鱼合称为我国的“四大家鱼”。鲤(Cyprinus),种类甚多,我国至少有20种,但以鲤鱼(Cyprinus carpio)为最常见,其他种类大多局限于云南省境内;口须2对;咽齿3行,臼齿状,鲤鱼是我国最早进行饲养的鱼种,现已成为世界性的重要养殖对象。鲫鱼(Carassius auratus),形似鲤鱼,无口须;咽齿一行,杂食性;适应性强于任何其他鲤科鱼类,可生活在江河、池塘、温泉以及含盐度高达4.5‰的草原、沙漠内陆湖泊中。早在宋朝就已有饲养鲫鱼的记载,古名鲋鱼,它不但是我国最普通的食用鱼类,而且还由此育成了许多金鱼品种。鳊鱼(Parabramis pekinensis),体侧扁,呈菱形,头和口均小,肛孔至胸鳍之间有腹棱;背鳍前有一大而光滑的硬棘;咽齿3行;广布全国各地,生活于水的中、下层,因主食植物性饵料,故又称草鳊。团头鲂(Megalobrama amblycephalua),形似鳊而大,腹棱局限于肛孔与腹鳍之间;栖于静水水域的中、下层,吃植物性饲料和少量浮游动物;主要分布于长江中游地区,有“武昌鱼”之称。红鳍鲌( Culter erythropterus)是鲌类中较常见的一种,体长,侧扁;下颌突出而上翘,口裂呈垂直状;咽齿3行;胸鳍下方至肛孔有发达的腹棱,为中、上层肉食性鱼类,以小鱼和无脊椎动物为主食;广布于全国,数量多,为中、小型经济鱼类。鲮(Cir-rhinus molitorella)体侧扁,口小,下位;口须2对;咽齿3行;胸鳍后上方8~9枚鳞片的基部有一黑斑;栖于南方水域的底层;广东地区在清代就已开始饲养鲮鱼,现为广东、广西和海南等省的重要养殖鱼类之一。雅鲁鱼(Leuciscus walickii),黑龙江和黄河中游地区的特有鱼类,也是冬季冰上作业的主要捕捞对象,体侧扁,腹部圆;口前位,无口须;咽齿2行,齿细长,顶端钩状;中、上层鱼类,喜栖于江河口及水流较缓、底质多石和水质清沏处,吃小鱼和昆虫等动物性饲料。青海湖裸鲤(Gymnocypris przewalskii)在青海湖周围地区称为“湟鱼”和无鳞鱼,除肩带部和臀鳍两侧有鳞外,全身裸露无鳞;头锥形,口呈马蹄形,无须;咽齿2行;背鳍棘发达,后缘有锯齿;背部灰褐色或黄褐色,体侧有不规则褐斑;是青海湖经济价值最大的鱼类。裸裂尻鱼(Schizopygopsis)是一些生活于山溪激流中的高原鱼类,垂直分布高度可达 4 000m左右;全身除臀鳍基部和肩部有少量鳞片外,均裸露无鳞;咽齿2行,口下位,下颌前缘角质锐利,以此刮食岩石上和水底的植物腐屑等,8~9月在底质为砾石的流水中产卵繁殖;我国产7种,主要分布于黄河和长江的上游、柴达木河和雅鲁藏布江等水系。

鳅科(Cobitidae)中、小型底栖鱼类。体大多呈圆筒形;口小,有口须 3~5对;咽齿1行,齿数甚多;鳔小,前端被骨囊所包。我国产 100余种,较常见的有:花鳅(Cobitis taenia),皮下有眼下刺;触须3对;体细长而侧扁,侧线不完全;背部及体侧各有10余个褐斑,生活在江边或湖岸的浅水处,觅食枝角类、藻类和植物碎屑,主要分布于长江以北地区。泥鳅(Misgurnus anguillica-datus),无眼下刺,触须5对;栖息在各地的小河和沟溪中,有钻泥习性,肠壁薄,密布血管,可营肠呼吸,对水中缺氧有很强的适应能力。
(2)鲇形目(Siluriformes):有韦伯氏器;口大齿利,口须 1~4对;咽骨有细齿;体表裸露或局部被骨板;通常有脂鳍,胸鳍和背鳍常有一强大的鳍棘。鲇形目中包括许多肉食性经济鱼类,我国产10科,主要有:
胡子鲇科(Clariidae)小型鲇类,背、臀鳍长,几与尾鳍相连,无鳍棘和脂鳍;口须4对;鳃腔内有辅助呼吸器官。常见的有胡子鲇(Claris fuscus),昼伏夜出,觅食动物性饵料。广西、广东、福建等地均有养殖。
鲇科(Siluridae)背鳍单个,无鳍棘,臀鳍长,与小形的尾鳍相连;体无鳞,富粘液腺;口须2~3对;肉食性,常危及池塘养鱼业,是清塘的主要对象。鲇(Silurus asotus)是本科中最常见的经济鱼类,栖于水域的中、下层,夜间外出觅食小鱼,除青藏高原和新疆外,全国各地均产。
5.银汉鱼总目(Atherinomorpha)体被圆鳞;腹鳍腹位,鳍条5~9枚,背鳍与臀鳍对生。主要目、种有(图16-39):
(1)鳉形目(Cyprinodontiformes):鳍无棘,背鳍一个,位于臀鳍上方,无侧线,为热带及亚热带淡水中的小型鱼类。我国产2种,即青鳉科(Oryziatidae)的青鳉(Oryzias latipes)和食蚊鱼科(Poeciliidae)的食蚊鱼(Gambusia affinis)。青鳉头宽而扁平,口小,口裂向上,有颌齿,眼大;栖于湖泊、小河、沟渠、池塘的浅水处,吃浮游动物和孑孓,也吞食养殖鱼的鱼卵和鱼苗,被列为清野的目标。食蚊鱼又名柳条鱼,原产于中、南美洲,素以嗜食孑孓著称,常被世界各国移殖用来消灭疟疾;卵胎生,每年繁殖3~5次,一次产仔鱼10~77尾。
(2)颌针鱼目(Beloniformes):鳍无棘,背鳍一个;侧线低位,与腹缘平行。大多是海洋鱼类,我国有鱼科(Hemirhamphidae)、飞鱼科(Exocoetidae)和颌针鱼科(Belonidae):鱼科的上颌短,下颌前伸成针状,两颌于相对部分具齿,常见种为鱼(Hemirhamphus sinensis),生活于近岸海区,是集群性上层鱼类。飞鱼科胸鳍高位,长大如翼,背鳍与臀鳍同形相对,尾鳍深叉而下叶较长;大多是热带和亚热带的浅海上层鱼类;喜集群,具趋光性,擅滑翔,可伸展胸鳍在海面停留十余秒钟,最长滑行距离达百米以上,常见而有经济价值的是尖头燕鳐鱼(Cypselurus oxycepha-lus)。颌针鱼科的两颌均延长如针状,外侧一列颌齿为稀疏的犬牙,我国沿海常见的是扁颌针鱼(Ablennes anastomella),活泼擅泳,为近海上层肉食性鱼类,肉味美。
6.鲑鲈总目(Parapercomorpha)体被圆鳞或皮肤裸露;许多种类颐部有一小须;腹鳍胸位或喉位,背鳍1~ 3个,臀鳍1~ 2个。(图 16-44)。江鳕(Lota lota)有背鳍2个,臀鳍1个,最大个体可长达1m而重25kg,日间隐伏在石缝洞穴中,夜间外出捕食鱼类,分布于黑龙江水系及新疆额尔齐斯河。大头鳕(Gadus macrocephalus)有背鳍 3个,臀鳍 2个,是黄渤海经济鱼类之一。


7.鲈形总目(Percomorpha)胸鳍大多为胸位或喉位,鳍通常有鳍棘;体常被栉鳞,但也偶有骨板或皮肤裸露的情况。本总目绝大多数为海鱼,种类极其繁多,共10目,常见种类有(图16-41):
(1)刺鱼目(Gasterosteiformes):吻大多呈管状,许多种类体被骨板,背鳍1~2个,有时第一背鳍为游离的棘组成。我国产7科,常见种有:刺鱼科(Gasterosteidae)的中华多刺鱼(Pungitissinensis),体侧扁,尾柄窄细;口大,两颌有锐齿;雄鱼具有善斗、以肾脏分泌物筑巢及卫护鱼卵的习性,是北方淡水中的冷水性小鱼。海龙科(Syngnatnidae),体长形,全身被环形骨板;吻呈长管状,口前位,鳃束状,鳃孔小;无腹鳍,尾鳍小或消失;雄鱼常由尾部下方或腹部皮褶形成育儿囊,容纳雌鱼产卵及受精卵发育,稚鱼孵化后,囊即张开,稚鱼游出体外;本科包括海龙(Syngnathus)和海马(Hippocampus)等近海小型鱼类,常栖于海藻丛中,吸食浮游甲壳动物;干制品为传统中药材。
(2)鲻形目(Mugiliformes):背鳍2个,前后分离,第一背鳍由鳍棘组成;腹鳍腹位或亚胸位。常见代表有:鲻科(Mugilidae),齿细小,绒毛状;鳃耙细密,形成滤器;第一背鳍由4枝鳍棘组成;分布于热带和亚热带海洋,有些种类也能进入咸淡水生活,我国产13种,常见种为鲻鱼(Mugilcephalus),体被栉鳞,口下位,具齿;脂眼睑发达;为近海中下层鱼类,以下颌刮食泥底表面的藻类和有机碎屑,是我国港养的主要对象,此外,与鲻鱼同为北方港养鱼类的还有梭鱼(Liza soiuy),脂眼睑不发达,眼稍带红色,4~6月为繁殖期。
(3)合鳃目(Synbranchiformes):体形似鳗,无胸鳍和腹鳍,奇鳍彼此相连,无鳍棘。左、右鳃孔位于头的腹面合而为一,鳃小而不发达,主要由咽腔内壁表皮代营呼吸作用。无鳔。我国只产合鳃科(Synbranchidae)的黄鳝(Monopterus albus),体光滑无鳞,全身黄褐色,满布不规则的黑色斑点。栖于泥质土穴或堤岸的裂隙内,夜间外出觅食;常将身体的前半部竖起露出水面,借咽腔进行呼吸。具有性逆转的特性。除青藏高原外,分布遍及全国,是江南地区常见的食用鱼类。
(4)鲈形目(Perciformes):腹鳍胸位或喉位,鳍条1~5枚;背鳍2个,分别由鳍棘和鳍条组成,无脂鳍;体大多被栉鳞。鳔无鳔管。本目为鱼纲中种类最多的一个目,其中包括许多重要的海产经济鱼种。我国产 85科,主要有(图 16-41):
科(Serranidae)下颌通常长于上颌,颌齿绒毛状或杂有犬牙;头和颊部被鳞,鳃盖骨上有棘。种类多,尤以鲈鱼(Lateolabrax japonicus)、鳜鱼( Siniperca chuatsi)和石斑鱼等优质经济鱼类最负盛名。
石首鱼科(Sciaenidae)头上有发达的粘液腔,颐部有粘液孔或小须;颌齿细小,间或有犬牙;耳石大。鳔的结构复杂,常有多对侧枝。本科为暖水性鱼类,繁殖季节常洄游到沿海浅滩产卵。我国产37种,在渔业捕捞中占有重要地位,经济价值最高的是大黄鱼(Pseudosciaena cro-cea)和小黄鱼(Pseudosciaena polyactis)同属我国四大海产经济鱼类而与带鱼、鳓鱼齐名。大黄鱼的尾柄长为尾柄高的3倍多,小黄鱼的尾柄短,仅为尾柄长的2倍多。近年来,这两种黄鱼的产量已大幅度下降,应注意资源保护。此外,体形较大而具犬牙的鱼(Miichthys miiuy)、体侧有波状斜纹的黄姑鱼(Nibea albiflora)和头背中央具有骨棱的梅童鱼(Collichthys lucidus)等也是常见的食用鱼类。
鲹科(Carangidae)体形不一,被圆鳞;尾柄细小;侧线完全,一部分或全部侧线鳞特化成棱鳞。第二背鳍及臀鳍后方常有一个或几个小鳍。尾鳍叉形。大、中型海鱼,善泳,大多生活于温暖的海洋上层,只有少数种类在夏季游向北方。分布广和产量大的有蓝圆鲹(Decapterus maru-adsi),和竹鱼(Trachurus japonicus)。
鲷科(Sparidae)体呈椭圆形;头大,口端位;齿强,两颌前端的齿呈犬牙状,两侧为臼齿或颗粒状牙。背鳍和臀鳍前方有沟,可容纳鳍棘藏于沟内。本科为温、热带海洋底层鱼类,我国产9种,常见的有:真鲷(Chrysophrys major),又名“加级鱼”,体淡红色,饰有宝石蓝色斑点,喜集群,游泳迅速,以贝类和甲壳动物为食,繁殖季节游向近岸,是北方名贵的食用鱼类。黑鲷( Sparusmacrocephalus)为浅海底层鱼类,体青灰色,带金属光泽,除胸鳍外,各鳍边缘均呈黑色,沿海各地都有分布。
?科(Echeneidae)第一背鳍特化成一长椭圆形吸盘,位于头顶部;第二背鳍及臀鳍无鳍棘,互相对生。本科为海洋上层鱼类,分布于热带和温带海域,常以其头部吸盘吸附在大鱼、海龟体上或船底,移迁远方。我国产 5种,常见种为?鱼(Echeneis naucrates)。
塘鳢科(Eletridae)体被栉鳞,无侧线。左右腹鳍在胸部彼此接近,但不愈合。尾鳍圆形。本科的常见种类有沙塘鳢(Odontobutis obscurus),体形粗壮,头大而宽扁;口上位,两颌具细齿;背面和体侧被栉鳞,腹面为圆鳞;体黑褐色而具黑斑,该鱼肉多刺少,美味可口,是长江以南地区的优质食用鱼。
带鱼科(Trichiuridae)体呈带状,尾端细如鞭梢。无鳞。有侧线。口大,下颌突出,颌齿大而税利。背鳍甚长;臀鳍由分离的短棘组成;胸鳍短小;腹鳍消失或退化成一对鳞片状突起。我国有 4种,以带鱼(Trichiurus haumela)为最常见,体银白色,布有黑色小点,尾暗黑色。性凶贪食,广泛摄食鱼类、软体动物和甲壳类,年捕获量居我国四大海鱼之首。
鲭科(Scombridae)体呈纺锤形,被小圆鳞。口大,两颌具细齿;鳃盖骨光滑无棘。背鳍2个,相距较远,第二背鳍与臀鳍同形并相对,鳍后均有分离小鳍;尾鳍深叉形,尾柄细长,基部两侧各有2~3条皮嵴。我国产17种,经济价值较大的是鲐鱼(Pnematophorus japonicus),背鳍和臀鳍后方各有5个小鳍,皮嵴2条,背侧青绿色,间有不规则的深蓝色彼纹。鲐鱼为暖水性远洋结群鱼类,是世界上产量最高的几种经济鱼类之一。青干金枪鱼(Thunnus tonggol)与狐鲣、鲣鱼、鲔鱼等统称金枪鱼,为大洋性洄游鱼类,小鳍8个,胸部鳞片大而形成“胸甲”。我国台湾东南及南海诸岛的海域内,均有丰富的金枪鱼资源有待于开发利用。
鲳科(Stromateidae)体侧扁而高,幼鱼的鳍棘发达,成鱼则埋于皮下而不显。吻圆钝,口、眼及鳃孔均小,颌齿细弱;食道的侧囊内有乳头状突起;体被小圆鳞,侧线完全;臀鳍与背鳍同形,无腹鳍,尾鳍叉形。本科为生活于外海的中下层鱼类,我国产2属3种,最常见的是银鲳(Stro-mateoides argenteus)。
攀鲈科(Anabantidae)体被栉鳞,有中断的侧线。有鳃上器作为辅助呼吸器官,是由第一鳃弓的咽鳃骨及上鳃骨扩大而形成的骨质瓣,其表面覆有一层粘膜并分布着大量血管,可用于直接从空气中摄取氧气。鳃盖骨有发达的锯齿。我国只产攀鲈(Anabas scandens)1种,分布于云南、广西、广东和海南等省的小河沟中,可经常离开栖居的水域,借助胸鳍和鳃盖锯齿的支撑在地表爬行,有时还能攀登上树,故有攀鲈之称。
斗鱼科(Belontiidae)头部被圆鳞,体侧为栉鳞,无侧线。具鳃上器。体褐色,有10余条深色横带,鳃盖后有一蓝色圆斑。我国淡水中产圆尾斗鱼(Macropodus opercularis)和叉尾斗鱼( M.chinensis),体态秀丽,有吐沫作巢的习性,雄鱼善斗,所以常被饲养供作观赏。
鳢科(Channidae)头似蛇形。鳍无棘。有鳃上器,在炎热干燥的季节,可因缺水钻进泥里呈蛰伏状态,靠气呼吸而生存。本科为底栖性的淡水鱼类,亲鱼有草筑巢、保护鱼卵和稚鱼的习性。我国产 2属 5种,常见的有乌鳢(Ophiocephalus argus),俗称黑鱼,有腹鳍,体黑绿色,布有蟒斑黑纹,性凶贪食,主要采用袭击方式猎食,是养殖业的大敌和清除目标。
刺鳅科(Mastacembelidae)体形如鳗,也像鳅类;被细鳞。背鳍、臀鳍与尾鳍相连;背鳍前方有 12~14枚游离的小棘;无腹鳍。我国产 2种,其中以刺鳅(Mastacembelus aculeatus)较为常见,臀鳍有3枚硬棘;无侧线;体侧有栅状横斑。
(5)鲉形目(Scorpaeniformes):第二眶下骨后延成一骨突,并与前鳃盖骨连接。头部粗壮,常具棘棱或骨板。胸鳍基底比较宽大。本目为一类广泛分布于热带、温带及寒带沿岸水域的海鱼,有些种类也进入河川、湖泊等淡水中。我国产11科,常见的有杜父鱼科(Cottidae)的松江鲈鱼( Trichidermis fasciatas)、鲉科(Scorpaenidae)的毒鲉、鲂鱼科(Trigilidae)的红娘鱼( Lepidotri-gla)和绿鳍鱼(Chelidoichthys)等。
松江鲈鱼的头部有棱无棘,口大,两颌和口盖上有绒毛状齿群;每边鳃盖膜上有2条橙红色斜纹,酷似鱼的鳃片,故有“四鳃鲈”之称,生活于近岸浅海,一般在与海相通的河川里生长肥育,冬季集群作降河洄游,到近海区进行繁殖,在贝壳内产卵。孵化后的幼鱼往淡水溯河上游,于江河中索食和生长。松江鲈鱼是我国四大淡水名鱼(黄河鲤鱼、兴凯湖鲌鱼、松花江鲑鱼)之一,也是很有价值的养殖对象。
(6)鲽形目(Pleuronectiformes):即通常所说的比目鱼类。体形侧扁,成鱼的眼、鼻、口、齿和偶鳍等均不对称,两侧的体色也各不相同,无眼侧通常颜色浅淡。背鳍及臀鳍的基底长,腹鳍胸位或喉位。肛门位置前移至胸鳍的后下方,且不在腹面正中。无鳔。幼鱼身体侧扁而左右对称,泳姿正常,变态后随同头骨的变化,一眼移位至另一侧,以无眼侧平卧水底营底栖生活,比目鱼即因此得名。本目是重要的海洋经济鱼类,我国产7科,种类较多,资源十分丰富,主要有:
鲆科(Bothidae)两眼均位于身体左侧。前鳃盖骨边缘游离。背鳍起点在眼的上方,有胸鳍,腹鳍不对称。常见种为牙鲆(Paralichthys olivaceus)。
鲽科(Pleuronectidae)双眼长在体的右侧。侧线于胸鳍上方大多无弓状弯曲。尾鳍圆形或略呈截形。本科常见种类有:高眼鲽(Cleisthenes herzensteini),上眼位于头部背缘的正中;有眼侧褐色,无斑纹。木叶鲽(Pleuronichthys cornutus),体呈卵圆形,头小;吻短;眼大而突出,眼间距甚窄,呈棱脊状;口小,有眼侧无齿,有眼侧有齿2~3行;广布于沿海各省。
鳎科(Soleidae)两眼均在体之右侧。口小,前位或下位;齿小,呈绒毛状;前鳃盖骨边缘不呈游离状。周身被小栉鳞,侧线直。背鳍、臀鳍与尾鳍相连。本科为生活于热带和亚热带海域的鲽形目鱼类,其中以条鲽(Zebria zebra)为最常见,有眼侧浅黄褐色,有平行的黑色横带,奇鳍边缘黑色。
舌鳎科(Cynoglossidae)形似鳎科而两眼位于身体左侧,有眼侧具侧线2~3条。无胸鳍。本科种类甚多,形状也极相似,常见种有:半滑舌鳎( Cynoglossus semilaevis),有眼侧褐色,被栉鳞,具侧线3条;无眼侧被圆鳞或杂有弱栉鳞;个体大,肉嫩味鲜,是优质的大型经济鱼类。
(7)形目(Tetrodontiformes):体短粗,皮肤裸露或被有刺、骨板、粒鳞等。上颌骨常与前颌骨愈合,齿锥形或门齿状,或愈合为喙状齿板。鳃孔小。腹鳍胸位或连同腰带骨一起消失。有些种类具气囊,能使胸腹部充气和膨胀,用以自卫或漂浮水面。大多为海洋鱼类,喜栖浅海海底或沿岸岩礁海区,只有少数种类有定居淡水或在一定季节进入江河的习性。我国产11科,主要科、种有:
革科(Aluteridae)体侧扁,尾柄细,被绒状小鳞。第一背鳍有鳍棘;左、右腹鳍各具一棘或合有一枚短棘。无气囊。本科约45种,大多分布于南海,常见和产值最大的是绿鳍马面(Navodon septentrionalis),因头形似马而得名,俗称橡皮鱼或扒皮鱼。口小,上颌齿2行,下颌齿单行。无侧线。腹鳍退化,合成一枚短棘。体蓝灰色,鳍呈蓝绿色。
箱科(Ostracidae)体短而高,包藏于具有3~5个棱脊的体甲内;尾部裸露,无鳞甲。背鳍一个,短小而无鳍棘;腹鳍消失。我国产4属8种,常见种类有体甲横断面呈三角形的驼背三棱箱(Rhinesomus gibbosus)、体甲为四棱状的粒突箱(Ostracion tuberculatus)及体甲四棱状而头背部具2枚棘突的角箱(O.cornutus)等,大多产于南海,均无经济价值。
科(Tetraodontidae)体短粗,皮肤光滑或被小刺。上、下颌分别愈合成 2个喙状齿板,有中央缝。背鳍单个,与臀鳍相似且对生,无鳍棘;缺乏腹鳍。有鳔,气囊发达。本科鱼类肉味细腻而美,但肝脏和生殖腺等均含有河豚毒素。类为近海底层肉食性鱼类,活动性差,少数可进入淡水中生活,我国产11属约38种,常见的有弓斑东方(Fugu ocellatus),俗称河和泡鱼,背部有1条暗色的鞍状横带,与胸鳍后上方的胸斑相连,背鳍基部也具一圆斑。生活于河口区,春季进入江河及淡水湖泊中。遇敌害时,气囊鼓胀,仰游漂浮在水面,体表小刺竖立,用以自卫。
翻车鱼科(Molidae)外形奇特,体侧扁而高,尾鳍短;背鳍和臀鳍均高,且同形相对,无腹鳍。无鳔及气囊。我国产2属2种,生活于热带海洋,以翻车鱼(Mola mola)较为常见。



 jpg
jpg

8.蟾鱼总目(Batrachoidomorpha)体短粗,平扁或侧扁,皮肤裸出,有小刺或小骨板。鳃孔小,位于胸鳍外侧的腹面。腹鳍胸位或喉位。本总目均为底栖的肉食性鱼类,以目(Lophi-iformes)较常见(图 16-42):
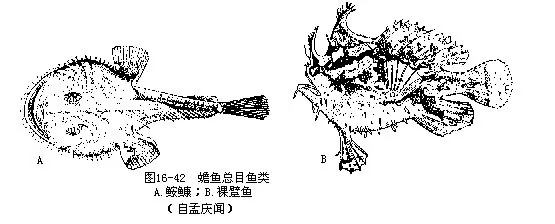
科(Lophiidae)体扁平;头大,口前位而宽阔;牙呈犬齿状,可向下倒伏。鳃孔大。第一背鳍的鳍棘分离,第一枚鳍棘位于吻上,末端呈肉质突起,用作诱捕其它鱼类为食;第二背鳍及臀鳍均位于后部。我国产黄(Lophius litulon)和黑(L.setigerus),分别分布于黄、渤海和南海、东海。
躄鱼科(Antennaridae)体侧扁;口小,上位,颌齿绒毛状。鳃孔小。第一背鳍有鳍棘 1~3枚,呈触手状。胸鳍具长形的假臂。如裸躄鱼(Histrio histrio),为暖温性低层鱼类,常栖息在海藻丛里,拟态于环境中,沿海均有分布。
第四节 鱼类的洄游
某些鱼类在生活史的各不同阶段,对生命活动的条件均有其特殊要求,因此必须有规律地在一定时期集成大群,沿着固定路线作长短距离不等的迁移,以转换生活环境的方式满足它们对生殖、索饵、越冬所要求的适宜条件,并在经过一段时期后又重返原地,鱼类的这种习性和行为叫作洄游(migration)。鱼类在由海入河的溯河洄游或自河至海的降河期间,都需要有一个转换调节渗透压机制的过程,以适应水质不同的环境改变。例如,鲥鱼、鲚鱼、鲑鱼、大麻哈鱼等都是溯河性洄游海鱼,它们在江河内产下的卵于淡水中进行孵化,幼鱼必须生长到鳃上的泌盐细胞充分发育和其它生理变化完成后,才能通过调节渗透压顺序渐进地回到海洋。这些鱼类及在淡水中生活一段时期后的幼鱼,于降河洄游移入海水生活的最初五日内,每天都要吞饮相当于体重4%~15%的海水,同时肾脏的滤泌作用也急剧地发生减弱,而泌盐细胞的泌盐机能和肾小管的再吸收作用却得到了加强,水分的排出也大大地减少,排水量仅为生活在淡水时的10%或更少。俟完全适应海水环境后,体液渗透压才由原来的高渗压转变成为低渗性溶液。
依据鱼类洄游的不同类型,可分为生殖洄游(breeding migration)、索食洄游(feeding migra-tion)和越冬洄游(overwintering migration)。它们三者间的关系如下图(图 16-43)。
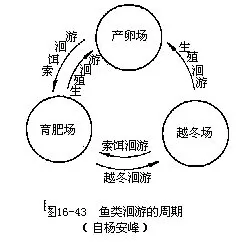
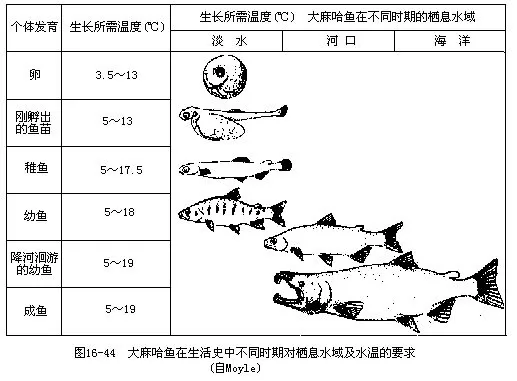
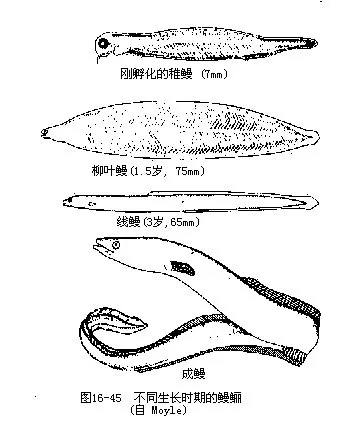
1.生殖洄游当鱼类生殖腺发育成熟时,脑垂体和性腺分泌的性激素对鱼体内部就会产生生理上的刺激,促使鱼类集合成群,为实现生殖目的而游向产卵场所,这种性质的迁徙称为生殖洄游。生殖洄游具有集群大、肥育程度高、游速快、停止进食和目的地远等特点。大多数海洋鱼类的生活史均在海洋中度过,它们的生殖洄游都是由远洋游向浅海,进行近海洄游。如小黄鱼、大黄鱼、带鱼、鳓鱼、鲷鱼、鲅鱼、鲐鱼、鳙鱼等。与此相反,青鱼、草鱼、鲢鱼、鳙鱼等是终生生活在江河中的淡水鱼类,它们的生殖洄游是从江河下游及其支流上溯到中、上游产卵,其行程可长达 1000~2 000km。此外,鲥鱼、大麻哈鱼、鲟鱼、鲚鱼、斑圆鲀、鱼、大银鱼等海鱼,在生殖季节成群溯河进入我国黑龙江、长江及其支流中产卵。繁殖活动结束后,大麻哈鱼因长途跋涉和饥疲交加终至体衰力竭而全部死亡,所以是一次性生殖的鱼类(图16-44)。其它鱼类产卵后,通常都在产卵场停留一段时期进行休整和肥育,然后循原路游回海中生活。产于我国的鳗鲡和松江鲈鱼是从淡水游向海洋去繁殖的仅有例子,这种洄游称为降河洄游。鳗鲡的性成熟年龄在8岁以上,集群游向深海进行繁殖的亲鳗于产卵后均因疲惫而死,无一生还。幼鳗孵化后,逐渐向亲鳗栖息的江河进行溯河洄游,此时的幼鳗周身透明,头细,体似柳叶状,故名柳叶鳗,经过生长和变态才成鳗形的线鳗(图16-45)。渔民在掌握了鳗鲡的周期性洄游规律后,每年初春于长江下游捕捞鳗鲡鱼苗进行养殖。
2.索饵洄游 鱼类为追踪捕食对象或寻觅饵料所进行的洄游,称作索饵洄游,例如我国福建南部蓝圆鲹(Decapterus maruadsi)追随犀鳕及带鱼追食隆头鱼类(拟隆头鱼 Pseudolabrus、海猪鱼Halichoeres)的集群洄游。索食洄游在结束繁殖期或接近性成熟的鱼群中表现得较明显而强烈,它们需要通过索食摄取和补充因生殖洄游和繁殖过程中所消耗的巨大能量,并且也为鱼类恢复体能、增强体质,以及积贮大量营养物以供生长、越冬和性腺再次发育的需要。有些鱼类(小黄鱼)于索饵洄游途中还伴有垂直洄游现象,它们在采食场对所追食的饵料生物作朝降暮升的垂直分布的移动。及时分析经济鱼类的主要饵料及其分布、移动、数量变化等基本情况,就能精确地预测鱼群的所在处和游动趋向,直接有利于渔业捕捞。
3.越冬洄游当秋季气温下降影响水温时,鱼类为寻求适宜水温常集结成群,从索饵的海区或湖泊中分别转移到越冬海区或江河深处,这种洄游叫作越冬洄游。鱼类进入越冬区后,即潜至水底或埋身淤泥内,体表被有一层粘液,暂时停止进食,很少活动,降低新陈代谢,以度过寒冷的冬季。
生殖洄游、索饵洄游和越冬洄游是鱼类生活周期中不可缺少的环节,但是三者又以各自的特点和不同目的而互相区别。洄游为鱼类创造最有利于繁殖、营养和越冬的条件,是保证鱼类维持生存和种族繁衍的适应行为,而这种适应是在长期进化过程中形成并由遗传性固定而成为本能的。至于诱发鱼类洄游和决定洄游路线的原因是极其复杂的,不仅与鱼类自身的生理状况有关,也与季节、温度、食源、海流、水质变化等都有直接或间接的关系,同时也与遗传性密切相关。研究鱼类洄游的规律,不但具有理论意义,而且在渔业生产上也有重大的经济价值。
第五节 鱼类的经济意义
一、鱼类资源利用
鱼类与人类的关系源远流长,考古学家于出土文物中多次发现过石质的鱼钩、鱼叉和鱼网坠,证实了早在人类发展初期曾经历过以捕鱼和狩猎为主要生产方式的渔猎时代,我国学者还在公元前11世纪的殷墟内,发现了青、草、鲤、赤眼鳟、黄颡和鲻等6种鱼的骨骸。
鱼类是水产事业的主体,具有突出的经济意义。渔业生产随同科学捕鱼和养鱼新技术的发展,渔获量连年得到增产,据联合国公布的资料,20世纪 80年代末,全世界的年渔产量已接近 1亿t,除30%用作饲料外,70%均为人类食品。我国于1991年的渔业产量创下了1320万t的新记录,这个数目比1951年的一百多万吨增长了10倍,并超过日本和原苏联而居世界各国之首,人均年食鱼消费量也由 1979年的 4.8kg提高到 11kg。
鱼的肉味鲜美,是高蛋白、低脂肪、高能量、易消化的优质食品,营养丰富,蛋白质含量16%~25%(带鱼16.3%,鲢17.3%,黄鳝18.0%,泥鳅18.43%,大黄鱼18.8%,乌鳢19.8%,鳜鱼 20.3%,鲤 23.9%),明显地高于牛奶、鸡蛋,与鸡肉、牛肉、羊肉和猪肉等(19.3%~20.3%)不相上下。此外鱼肉中还有人类必需和容易吸收的脂肪、钙、磷、铁、赖氨酸、硫胺素、核黄素、尼克酸、抗坏血酸和多种维生素。
除鲜食和加工成“海洋牛肉”(肉组织状的浓缩鱼蛋白)、鱼翅(鲨鱼鳍)、鱼肚(鱼鳔)和鱼唇(鲨、鳐的吻软骨)等珍馐外,渔产品还被开发进行了广泛的综合利用,为工业和医药生产提供原料。鱼鳞可提取和制成鱼光鳞、鱼鳞胶、盐酸鸟粪素、咖啡因、黄嘌呤、鳞酱油、磷酸钙肥料等。鲨鱼和鲀类的皮可做成上等皮革制品,鱼皮的优点是容易染色,产品成本也低,只及蛇皮的1/10,因此很受消费者的欢迎。鱼类内脏器官的利用是多方面的:含有高脂率的鱼肝(鳕、鲨、鲆、鲽等)可提制鱼肝油,是医疗及防治人体疾病的常用营养药物;精巢可制鱼精蛋白;鱼胰可提取胰岛素,其中以鲣鱼和鲔鱼生产的胰岛素质量最佳;鱼胆是提炼胆色素钙盐的原料,可用作细菌培养剂的胆盐和制造人造牛黄。由深海鱼类压榨取得的鱼油含有高度不饱和脂肪酸,医药上可用于减少人体血液中的胆甾醇;从鱼油中提取的二十碳五烯酸(EPA),可制成防治高血压形成的脑血栓的新药—血液凝固缓和剂;通过处理的鱼油,能制成一种具有特殊风味和稳定性良好的凝固脂肪,用作生产人造鱼黄油;制取氨基甲酸乙脂和环氧树脂,作为涂料;用鱼油处理皮革,可使之赋予黄色、在矿石浮选中用以分离低价铁等、作为润滑剂和防水剂。鲀类的血液和内脏含有河鲀毒素(Tetrodinine),有抑制人血中胆碱酯酶活力的作用,能麻痹末梢神经和中枢神经,以此制成的河鲀毒素针剂可治疗痉挛、外伤疼痛、神经痛及晚期癌症患者的止痛。海马和海龙是传统中药,有滋补、安神、散结、舒筋活络等功效。
鱼的头、骨、刺等废弃物和不堪食用的杂鱼,常用于生产鱼粉,或采用生物发酵制造液化饲料。鱼粉是有些发达国家养猪和养禽业增产所不可缺少的填加剂,丹麦的渔获量中约有2/3是用以加工成鱼粉作为家禽家畜饲料的。
食蚊鱼、鳟鱼、斗鱼、麦穗鱼、棒花鱼(Abbottina rivularis)和黄颡鱼等小型鱼类都能大量吞食孑孓,是蚊子幼虫的天敌,对其数量控制和防止由蚊类传播的脑炎、黄热病、疟疾和血丝虫病等都有积极的作用,间接有益于人类。
二、海洋渔业
我国地处太平洋的西北岸,海疆辽阔,北起渤海的辽东湾,南至北纬4°附近的曾母暗沙,跨越了热带、亚热带和温带3个气候区。我国的海域东面连接太平洋,西南面通过马六甲海峡和爪哇海,与印度洋相通,自北向南包括渤海、黄海、东海和南海等四大海区。海岸线长约11 000km,连同沿海的 5 000多个岛屿在内,海岸线总长约 23 000km,超过地球圆周长度的一半。我国沿海有两种不同反向海流,一是由南向北流动的暖流,水温高而盐分大,一是自北往南流向的寒流,水温低,盐分少。这两种海流的交汇处,涌升流发达,经常形成大量浮游生物集中并引来各种鱼类在此索食;来自不同海流中的鱼类由于水温和含盐量的骤变,出现暂时滞留于该地区的现象,从而使海流交汇地带成为很好的渔场。国内著名的渔场有渤海、大沙、舟山、粤西、粤东和北海湾等,渔场主要位于水深 200m以内的大陆架,面积约 80.75万km2,相当于 1.5亿ha水面,约为全世界浅海区总面积的23.7%,是最适宜海鱼生息及进行拖网作业生产的区域。港湾和滩涂水面约100万ha,可供开展水产养殖的有49万ha,而适于鱼类养殖的约占13多万ha。
海洋沿岸有长江、黄河、珠江等许多河流倾注入海,不断带来大量无机盐类和有机物质,因此海水中的营养物质含量高,水质肥沃,适于各类海洋生物和鱼类的栖息、生长和繁殖,是经济鱼类生长和繁殖的良好场所。我国的海洋鱼类资源十分丰富,约有2 000种左右,占全国所产鱼种的3/4,其中经济价值最大的有带鱼、小黄鱼、大黄鱼、鳓鱼过去号称四大海鱼,总产量为全国海洋水产品的1/4。此外,产量较大的还有鲥鱼、青鳞鱼、石斑鱼、鱼、大眼鲷、蓝圆鲹、竹鱼、红鳍笛鲷、鲱鲤、金枪鱼、扁舵鲣、鲔鱼、太平洋鲱鱼、金色小沙丁鱼、斑、鳀鱼、黄鲫、龙头鱼、蛇鲻、海鳗、大头鳕、鲻鱼、梭鱼、真鲷、黄鲷、二长棘鲷、鲈鱼、鱼、黄姑鱼、鲐鱼、马鲛鱼、鲳鱼、红娘鱼、牙鲆、木叶鲽、高眼鲽、舌鳎、东方鲀、海鲇、鲨类、鳐类等200多种。鲱形目和鳕形目鱼类的捕获量分别为世界渔业生产的第一二位,占年生产总量的1/5~1/4,但是这两目鱼类的产量在我国的海洋渔业中并不占有重要位置。
随着我国人民生活水平不断提高,对于水产品和蛋白质的需要正在与日俱增,从而促进了渔业生产的快速增长,海洋捕捞在增殖、保护近海鱼类资源的基础上,开始向远洋渔业生产发展,并且在1986年达到年捕海鱼390万吨的历史最高记录。
旧中国长期处于内向型的海洋渔业格局,生产水平低,20世纪40年代末,年产量曾跌落到35万 t的低谷,比 1936年还减少了 100多万 t。中华人民共和国成立后,渔获量回升至 100万t,到20世纪60年代中期达到200万t,70年代增加到300万t,直到20世纪80年代末才登上400万t左右的台阶。内向型海洋渔业的基本特征就是完全依靠开发利用本国邻近海域的鱼类资源,其中又以近海资源为主。直到1984年,近海渔业产量仍占海洋渔业总产量的90%以上。我国是世界三大渔业国之一,但是远洋渔业于1985年才刚刚开始起步,当时的年产量仅10万t,与前苏联年产量400多万t及日本年产量200多万t的差距甚大,就是与韩国年产量80多万t,我国台湾省的60万t相比,也还有不少差距。当前,人民对渔产品需要的日益增长和近海渔业资源衰退的矛盾逐渐加剧,走出近海、减轻近海的捕捞强度及发展远洋渔业已是我国渔业发展的必然趋势,也是决定我国海洋捕捞业前途兴衰的关键。
据联合国粮农组织渔业委员会预测,到本世末,世界海洋的年捕鱼量将由现在的1亿t增加到1.3亿t,而潜在的年产量则可能为2.4亿~2.5亿t。因此,目前世界上还有一些海区的渔业资源蕴藏着开发潜力,例如,太平洋北部的狭鳕(Theragca chalcogramma)资源相当丰富,每年的可捕量约1000多万t,然而现在的年开发量仅600万t;太平洋西北部的拟沙丁鱼(Sardinopsmelanostictus)渔场;西非大西洋的拟牙(Pseudotolithus)、赤(Senegalensis)、海鲇(Ariusparkeri)、十指马鲅(Galeoides decadactylus)、带鱼(Trichiurus lepturus)、裘氏石鲈(Pomadosysjubelini)、非洲鸡笼鲳(Dreeane africana)等中、上层鱼类资源;南太平洋、印度洋的金枪鱼类资源等,都具有进行合作开发的美好前景。中国水产总公司于1985年起先后组织了一批远洋渔业船队,分赴西非、北太平洋、南美、印度洋、南太平洋及东南亚等渔业海域,采用拖网和延钓作业开展捕鱼。我国在5年的远洋捕鱼期间累计捕鱼量已达到47万多t,并初步建成一支既能抓生产,又能抓经营管理;既有适于公海大洋性生产的大型渔轮,也有适应大陆架生产的中小型渔轮;既能进行中底层拖网作业,也能进行延钓金枪鱼作业;既有生产渔船,又有配套的加工运输船的中国远洋渔业船队。
海水鱼类养殖是海洋渔业的一部分,即咸淡水养殖,又称港塭养鱼。这是利用沿海港湾、海汊或海、淡水交汇的滩涂及低洼地带,加以人工筑堤围港,开沟建闸,贮蓄海水,利用潮汐的涨退纳入鱼苗进行养殖的一项生产事业。这种养殖方式,在我国北方沿海各省称为港养,福建叫海埭养殖,广东叫鱼埭养殖。我国有港湾和滩涂面积约 150万ha,适于海鱼养殖的就有13多万ha。我国港塭养鱼的历史比较悠久,早在明、清两代已有关于论述海鱼养殖的专著,黄省曾在《养鱼经》中记载:“鲻鱼,松之人于潮泥地凿池。仲春潮水中捕盈寸者养之。秋而盈尺。背腹皆腴。为池鱼之最。是食泥。与百药无忌。”由此可见当时在海滩凿池养殖鲻鱼是颇有成就的。胡世安在《异国賛闰集》里写道:“流鱼,如水中之花,喘喘而至。视之几不辨,乃鱼苗也。谚云:‘正乌二鲈’。正月收而放之池,皆为鲻鱼。过二月则鲈半之。鲈食鱼,蓄鱼者呼为鱼虎。故多于正月收种。其细似海虾,如谷苗,植之而大。流鱼正苗时也。”该书集中概括了当时渔民开展海鱼养殖的生产经验。“正乌二鲈”表明渔民收集的鱼苗中既有鲻鱼,也有鲈鱼苗,同时又指出“鲈食鱼,蓄鱼者呼为鱼虎”,对鱼、虾养殖的危害很大。渔民们利用鲻、鲈鱼苗出现的时间差异,在正月及时收集和放养鲻鱼苗,这是防止在养鲻池内混进鲈鱼苗的有效办法。这些经验至今仍在海水养殖鲻鱼方面起着指导生产的作用。
国外开展港养业较早的是印度尼西亚、菲律宾和泰国,他们利用沿海沼泽地带进行鱼、虾混养,尤其是鲻鱼、遮目鱼等与虾类混养。70年代起,日本的港湾养鱼有了很大发展,除了传统的利用废盐田养殖外,还使用尼龙网拦网进行海港养殖,以及利用电场控制鱼群。目前,网箱养殖在世界各国的沿海地区相当普遍,其优点是投资少,能充分利用水体和饵料,养殖密度大,单位面积产量高。
我国沿海有许多港湾是港塭养鱼的良好场地,但已被利用的还只有1.3万ha,主要集中于河北、山东和广东等省,其中以广东省的养殖面积最大。当前港塭的主要生产方式是引水纳苗,比较粗放,引进的放养鱼苗中既有吃植物性饵料的鲻鱼、梭鱼、斑等,也有鲈鱼、鲷鱼及虎鱼等肉食性鱼类,鱼、虾类之间的残食现象相当严重,直接影响了港塭养鱼的产量。由于很多港塭的自然条件优越,面积广大,养殖方法简便,投入的人力、物力不多,生产周期短,每年可以提供数量可观的鱼虾,且鱼货新鲜,随取随有,故有活鱼仓之称。在不断开拓海鱼养殖面积的同时,若能着重解决养殖中的优种鱼苗的来源、清除野鱼、施肥和防止污染等问题,我国的港塭养鱼事业,前途将是不可估量的。
三、淡水渔业
我国内陆水面的面积约2 000万公顷,可作为渔业生产的淡水水域约 700万公顷,还有1000多万公顷稻田和浅水荡滩也可用于结合养鱼。我国的淡水渔业资源丰富,有淡水鱼种800种左右,具经济价值的就有250多种,已发展成(包括引进鱼种)养殖对象的已有青、草、鲢、鳙、鲤、鲫、鳊、鲂、鲴、鳗、黄鳝等70多种。
中国是淡水养鱼事业发展最早的国家,有人根据在殷墟出土的甲骨卜辞有“贞其雨,在圃渔”的记载,考证出人们在公元前 1140多年的殷商时代已开始了池塘饲养鲤鱼。公元前460年,陶朱公范蠡在综合养鱼的实践基础上,撰写了《养鱼经》,这是世界上留传至今最古老的养鱼文献,遗憾的是原书已经失传,现在所依据的是后魏夏思勰所编《齐民要术》中的一段内容,全文共343字。池塘养鲤在唐朝逐渐衰落,原因是鲤与皇族的“李”姓谐音,故下令禁止捕食,渔民被迫只能到江湖中去捕捞鲢、鳙、青、草、鲮等鱼苗,作为新的池塘养殖对象,并开始在稻田内试养草鱼,从而使池塘养鱼由单养一种鲤鱼,发展成多种鱼类的混养,这是养殖技术上的跃进和渔民的卓越创造。宋代周密的《癸辛杂识》(约1243年)中有渔民在运输鱼苗到达目的地后用布兜培育的叙述,这一措施可以看作是我国网箱养鱼的前身。明朝的淡水养鱼技术有了较大发展,不论在黄省曾的《养鱼经》还是徐光启所著的《农政全书》内,都有关于鱼池建造、鱼种搭配、放鱼数量、饵料投喂、饲养方式、鱼病防治的记载。由此可见,池塘饲养青、草、鲢、鳙四种鱼类的技术在当时已经比较完整和渐趋成熟,为近代池塘养鱼事业的发展奠定了坚实基础。
长期以来,池塘养鱼一直是我国淡水渔业生产的主体和支柱,大水面放养和天然捕捞不超过淡水渔业总产量的20%。20世纪下半叶,池塘养殖在科学技术的指导下,得到很大发展。1958年,在全国普遍开展养鱼的形势下,以江苏省无锡市河埒乡为典型,并归纳各地渔民的经验总结成“八字精养法”(或称“八字养鱼经”),即池塘养鱼必需加强水(水源条件)、种(苗足质优)、饵(充分质鲜)、混(混合放养)、密(合理密养)、轮(轮捕)、防(病害防治)、管(科学管理)等8个方面的管理工作。水、种、饵是养鱼的物质基础,其它方面则是发挥其增产潜力的技术措施,八字精养法在国内推广和全面贯彻后,极大地促进了池塘养鱼的高产稳产。
合理密放和混养技术是我国渔民的创举。这是一种设计与调整养殖鱼种类、性比、龄期组成结构,实行多层次、多周期生产的立体养殖体系。即根据各种养殖鱼的食性(滤食性的鲢鳙类、草食性的鲂草类、肉食性的鲤青类)、栖息水层(上层鱼、中上层鱼、中下层鱼、底层鱼)和生长周期不同,按一定比例的数量进行混养,以保持平衡,达到在空间上、物质上和能量上可以多层次分级利用养殖水体的栖息位、营养位和市场位。合理密放和混合放养既有利于鱼类充分利用水体空间和饵料资源,提高鱼的生长率及群体产量,又可调剂市场的鲜活鱼供应,加速资金周转,增加经济效益。
1958年,广东水产研究所首先采用注射鲤鱼脑垂体摧青和模拟天然流水刺激相结合的方法,使鱼的生态因素与生理因素结合起来,诱导池养鲢、鳙产卵成功并孵出鱼苗。此后,草、青、鲮、鳜、黄鳝、松江鲈、中华倒刺鲃、长吻、泥鳅、胭脂鱼、鲥鱼、香鱼、中华鲟、短吻银鱼、花、大口鲶、黄颡鱼等,也相继获得人工繁殖成功。这就使我国的养殖鱼类的鱼苗来源彻底改变了过去长期依赖天然捕捞的被动局面和养殖品种单调的状况,可以充分满足养鱼生产的需要。
随着各国工业化的发展,全球性的水资源短缺现象日趋明显,世界上已有 40%的河流不同程度地被污染。省水型的工业化流水养鱼业于19世纪末在美、日等少数工业国开始兴起,将养殖鱼高密度地投放在小面积的养鱼工厂中,进行科学化饲养,把每亩水面的鱼产量提高了几十倍、几百倍,使养鱼业进入一个崭新的发展阶段。为解决城市养鱼的水源及能源问题,我国从 80年代起开展了利用地热和工厂的余热水进行人工流水高密度养鱼。养鱼工厂不仅节约了鱼池面积和水源,并且可以不受自然条件的影响,鱼的生长快,成活率高,产量大,经济效益可观,为缓解城市供鱼具有积极意义。
我国学者以遗传学原理为指导,用人工杂交的方法,通过种内杂交为主、远缘杂交为辅的途径,育成了一些具有优良性能和抗病力强的新型养殖鱼种。例如 1975年,水生生物研究所利用诱导雌核发育,由兴国红鲤与方正银鲫杂交培育成异育银鲫,由于精子没有参加受精过程,故得此鱼名,异育银鲫的生长要比普通鲫鱼快2~3倍;1980年又采用传代体细胞获得我国第一尾无性生殖的鲫鱼; 1985年,把一种生长基因注射到泥鳅的受精卵内,孵出的转基因泥鳅比未经注射的个体生长速度快3~4倍。淡水渔业研究中心于 1982—1987年也获得培育转基因鲤鱼、鲫鱼和团头鲂成功。此外,还开展了辐射育种的工作,探索了运用α、β、γ射线,快中子,慢中子,超声波等诱发突变,为缩短鱼苗培育周期和培养大规格鱼种提供科学依据。
1990年,我国淡水渔业的总产量已达524万t,超过世界内陆水域渔业总产量的40%而名列世界各国首位。淡水渔业的产量中有446万t来自池塘养鱼,这个数量几乎是40年前年产量的30倍,而亩单产量在很多地区已达到0.5t,少数鱼池甚至还有超过1t的记录,由此可见池塘养鱼在淡水渔业生产中地位的重要性。预计在对鱼类进行深入研究和在新科技的指导下,淡水渔业将得到更广泛的开发利用,并继续为促进我国淡水渔业的发展作出更大的贡献。
复习题
1.鱼类在脊椎动物中是最适于水生生活的一大类群。试从它们的形态结构上加以说明。
2.鱼纲和圆口纲有何异同点?这些异同点说明什么?
3.鱼的鳞、鳍和尾有哪些类型?
4.鱼类的骨骼系统有些什么特点?
5.鱼类消化道的结构和它们的食性有什么关系?
6.鳔的作用是什么?
7.列举鱼类循环系统的特点。
8.简述鱼类肾脏在调节体内渗透压方面所起的作用。
9.鱼类的视觉器和听觉平衡器的基本结构如何?
10.鱼类的脑和脊髓的基本结构和功能。
11.举例说明鱼类的两性异形。
12.概述鱼卵受精和发育的几种类型。
13.列举软骨鱼系和硬骨鱼系的特征。
14.软骨鱼系和硬骨鱼系各有哪些亚纲及总目?试举出1~3个实例。
15.小结鱼类的经济价值。
16.什么是鱼类的洄游?可分为几种类型?研究洄游有什么实际意义?
17.举例说明渔业生产与基础理论研究的关系。