�������и�����������ʢ��һʱ��������������䷱�࣬�ִ�����ֻ���������ꡢ������ߵȶ�����и��������Dz��������ϲ���������Python reticulatus����ȫ���ɴ�9.8m������С������Ϊ�ߵ�Բֺ����Sphaerodactylus argus����ȫ��ȴֻ��36mm�����ϼ������⣬�ֲ������鼰ȫ��������ϰ���������Ϊ���࣬����Ϣ��ƽԭ��ɽ�ء�ɭ�֡���ԭ����Į���������½ˮ��ȸ���������������������棨����ɳ��Phrynocephalus theobaldi����βɳ��Phrynocephalus eryth-rurus������ߴ�ֱ�ֲ���ɴﺣ����ǧ������ظ�ԭ��Į�ش���
��һ�� ���иٵ���Ҫ����
��
һ����Ĥ�Ѽ����ڶ����ݻ�ʷ�ϵ�����
��
�����ڼ������ˮ��½���������������У��������ϵͳ�����ڻ��ȫ�淢չ�Ļ����ϣ������ܶ��������Ĥ��ת������ж������Ĥ����һ��������Ҫ�ķ�Ծ��������Ĥ�ѵĽṹ�ͷ����ص㣬ʹ��Ĥ���ﳹ�װ����������ڸ��巢�����ڶ�ˮ����������ſ���ȷ����������½���Ͻ��з�ֳ��
�������ж��������Ĥ��Ϊ�˻��ѣ������ѻ�Ĥ��ȱ������ˮ�з������ڽ�Ĥ���⽺Ĥ������������������ѹܱ������ں��γɵĵ��ס������Ĥ��shell membrane�����ѿǣ�shell�����ѿǼ��ͣ���ʯ���ʻ���ά�ʹ��ɣ���ά���ѵ���״����������ˮ�������������е���˺ͷ�ֹ��ԭ�����롣�ѿDZ���������С�ף�ͨ�������ã��ɱ�֤��̥����ʱ���������л��������һ���ܴ���ѻ��ң����зḻ���ѻƣ�Ϊ�����ڼ����̥����Ӫ�����ʡ�
������̥�ڷ����ڼ䣬������Ĥ��amnion������ëĤ��chorion�������ң�allantois����һϵ����Ĥ����Ĥ���ﹲ�е����ԣ�Ҳ�DZ�֤��Ĥ��������½������ɷ�������Ҫ��Ӧ����Ĥ�ѵ���̥������ԭ���ں���������Χ��������¡��Ļ�״����——��Ĥ��ëĤ�ޣ����������Ļ�״���������������м��£���˴����Ϻʹ�ͨ���ΪΧ����������̥��2��Ĥ�����ڲ����Ĥ��������ëĤ������֮����һ�������������ǻ��exocoelom������Ĥ����̥��Χ�ڷ�յ���Ĥǻ��amnioticcavity���ڣ�ǻ�ڳ�����ˮ��ʹ��̥���������������һ��ˮ���н��з���������Ч�ط�ֹ�������������ˡ���ëĤ�����ڿ�Ĥ���档��̥���γ���Ĥ����ëĤ��ͬʱ����������������һ���䵱��������й�����٣���Ϊ���ҡ�����λ��������ǻ�ڣ���ڽ�����ëĤ������������ëĤ�ڱ��ϸ���ëϸѪ�ܣ���̥��ͨ����Ŀ�Ĥ���ѿǣ�ͬ���������彻�������⣬���һ���Ϊһ������ʢ����̥�³´�л�����������ᣨͼ18-1����
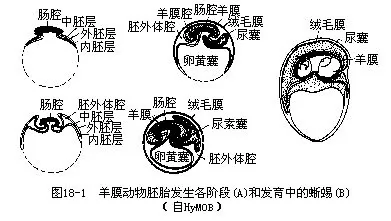
��
�������ж����ò���Ĥ�ѵ����Ժ����赽ˮ�з�ֳ��Ϊ���ж���ͨ��������Ӧ��ɺ������ֲ��������µ������������������
����3����ǰ�������иߵȺͽ����������������ж����ڵ����ϳ��ֺܿ�͵õ�����ķ�չ����Ϊ�ڵ����ϸ�����̬������ռ������λ�Ķ��Ȼ�������ж����л��ṹ�����Ƴ̶Ȳ�δ�ﵽ����緢չ�Ķ��壬����ߵȵ���ٺ��ٱȽϣ����ǻ��������Ϊ�ͼ�֮����������ж����뼹�����е�����Ĥ��һ���������ڱ��¶��������������������������٣��Լ����µ��ڻ��ܵIJ����ƣ�������ά�ֺ㶨�����£��ںܴ�̶���Ҫ�ܵ�������Ӱ�죻���Dz��������ڹ��ͻ���ߵ��¶Ȼ����У�������Ϻ�����Ķ��������ȸɺ����ļ�����Ҫ�������ߡ�
��
�������иٵ�����ṹ
��
������һ������ ���ж������������Ƭ���������β����������ɷ�Ϊ������̬�������ͣ����桢����Ш����ȣ����ػ���̬�����ͣ��ߺ�����ȣ�������ͣ���ͱ�ȣ����ֱ���Ӧ�ڵ��ܡ����ܡ�ˮ�ܺ�Ѩ�ӵȲ�ͬ���ʽ�������������⣬��������Ե�����Ϊͷ���������ɺ�β�����л�Ե��������ڻ����������⣩����Ĥ������������������֫ǿ��������ǰ��֫��Ϊ��ָ��ֺ����ĩ�˾�צ���������������ۺ��ھ���йֳ�����ѣ������꣩�����ѣ����桢�ߣ���Բ�Σ��ꡢ���β����Ϊ�ִ�������ϸ������β�ҡ�
����������Ƥ�� ���ж����Ƥ���ص��DZ�Ƥ�߶Ƚ��ʻ���Լ��10��ϸ�����ɣ����ⱻ�����ۣ�horny scale���������������۱���������Ч�ط�ֹ����ˮ�ֵ������������۵ı˴����ڲ��ֽ��ʲ�䱡��ʹ��Ƭ���ܻ�����ܻ��������������࣬���������������ۣ�keeled scale���������ڽ���������������Ƥ�����Ĺǰ��Ĥ�ߣ�abdominal ribs����������ɱ�Ƥ�γɵĽ��ʶܣ�hornyepidermal shield������Ƥ��Դ�Ĺǰ壨bony plate����ͬ�������ɡ�Ƥ�����Ƥ���ٺܲ�����ϳ�����Ƥ������λ�ڲ��ᡢ���ᡢ̥���ᡢ��Ƥ��ͱڻ�����������ڲ��йֳ��ǰ�Ĺ��٣�fem-oral gland�����ٿ����г��У���Ϊ�����ѣ�igunal pore�������ѣ�femoral pore����ǰ�ѣ�preanal pore���ȣ���������ڷ�ֳ�ڳ������ڿ��⣬�γɻ�ɫ�Ķ̴�״Сͻ�������ڽ���ʱ�ѳִ��ᣨͼ18-2��������йֳǻ�ڼ���β�Ļ������Լ��������ºͿ�䶼����һ�Գ��٣�scent gland�������ķ�������������ζ���������Դ��������Ի�ѭ��ζԴ�ҵ����������ż���䡣
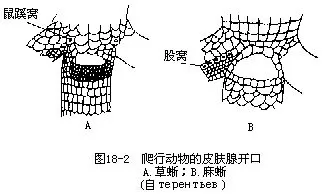
��
�������ж�����۱��ж��ڸ������ɣ���Ϊ��Ƥ��ecdysis����Ȼ����һ���е���Ƥ��������Ƥ��ʽ���ֶ��졣����Ѹ�ٵ�����ÿ������������Ƥһ�Σ������ߵ���Ƭ��������Ƥͻ��Ķ������ֲ�����Ƥ�������ӣ������ȥ�ɻ��µ���Ƥ�����У���ø�������£����ɵı�Ƥϸ����Ļ����ܽ����ʹ�ɵı�Ƥ���ʲ���������ϸ�����Ϸ������γɵ���ϸ������룬����Ƥ���������¡�
�����������ƤΪ��Ƭ���䣬�����������ԣ��ꡢ��������Ƥ���ɣ�ͨ�����Բ��ϸ�����ɵķ�ʽ���еġ�
������Ƥ�����ܵ���ά�����֯���ɣ��ں��ḻ��ɫ��ϸ��������ɫ�ؿ��������ڱ�Ƥ���ʲ㣩���ɴ����ɫ�����İ���ͼ����ͼ18-3�����������棨���ᡢ���ᡢʯ���ӡ�ɳ��ȣ���ɫ��ϸ����ֲ�������Դ�����ڷ����ٵĵ����£���������չ���������Ӷ�������ɫ�仯�������������ڿ��ٱ�ɫ������“��ɫ��”֮�ơ�����ı�ɫ������ʹ�����뻷����Ϊһɫ�ı��������⣬���������յر������ȼ����µĹ��ܡ�
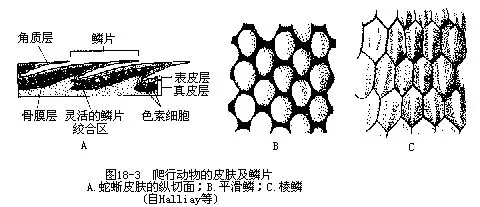
��
��������������ϵͳ ���ж���Ĺ���ϵͳ�����Ӳ�ǹ��ɣ������Ĺǻ��̶ȸߣ����ٱ������Dz��֡�
����1.�������
������1���ǽϸ߶�¡��Ϊ���ͣ�tropibasic type��������ǻ��չ�����������������������ǵ����ǻ��Ǻ�Ĥ����Ŀ����½���������������ġ�
������2���е�ö���������һö��������
������3������ǰ�Ǻ����ǵ���ͻ����ǵȵ�ͻ��ͬ�ϳɳ��͵Ĵ�����secondary pala-ta����ʹ��ǻ�е��ڱǿ�λ�ú��ơ�������Ľṹ����������Ϊ�������Ӧ��ˮ�в�ʳ���ػ�����ͼ18-4����

��
������4�����������ۿ�����1��2���ף�temporal fossa������������ԭ������ҧ�������ڵ������γɵĿ�����ҧ��������ʱ����ʹ���ļ�������������������䡣ͨ������λ�ڿ���ǡ�ȧ�ǡ��۹ǡ�����ǺͶ���֮�䣬���ϡ��µĹǹ��ֱ��Ϊ������������������������λ�ã��ɽ����ж����Ϊ�����ࣨanapsida���������ࣨparapsida���������ࣨsynap-sida����˫���ࣨdiapsida����4�����͡�����������������ǹ����ж����ԭʼ�����ͣ��ִ溣����Ҳȱ���ף������Եı仯�������ࡢ�������˫��������������ݱ���������������ֻ�е������ף����ɿ���Ǻ��۹ǹ��ɣ��������������ǵ����ж����������ࣨIchthyosauria���ͺ��������һ����е����ף�������ǡ��۹Ǻ�ȧ����Χ���Կ���Ǻ��۹ǹ������������ж����е����ࣨTheriodontia����֮���ִ�����ٶ��T�ɴ��ݽ�������˫���������������ϡ���2���ף��ִ���������桢�ߵ����ж�����Ǿ������࣬Ȼ���ڽ��������ж������˲�ͬ�̶ȵ��ػ���ֻ��Ш���ᣨSphenodon������������ŵ��͵�˫������ǡ������������ή�����˻������ܹ�����������������µ������ף������������˻�����ǵĻ����ϣ��ֽ�һ��ʧȥ����Ǽ�ȧ�ǣ��Ӷ���ȫ��ʧ����Ϊ�ۺ��ϡ����ֽ�Ĺǹ���ʹ���ߺϳ�һ����ף�ͼ18-5����
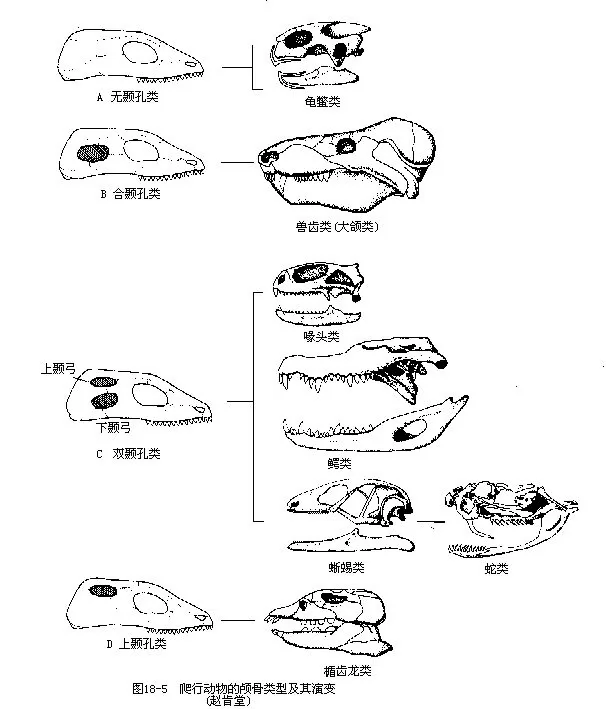
��
������5���ܶ�������������֮����б���Ƭ�γɵĿ������interorbital septum����
������6�������˹ؽڹ��⣬���гݹǡ��а�ǣ�splenial������ǣ�angular��������ǣ�surpra angular����״�ǣ�Coronoid���ȶ��Ĥ�ɹDz�����ɡ�
������7�������ѷֻ���½�ܼ����ﹲ�еľ���������������β��5����������2ö������һ���ڶ����ֱ��ػ�Ϊ���atlas��������axis��������Ŀ�����༰�����������ι����ӣ���ǿ�˺�֫�������ظ��ɡ��ǰ�����ǵ��������������ij�ͻ����������ɿɶ����ᣬʹͷ����ø��������ԣ��Ӷ�ʹͷ�����������˶�������ת����ͼ18-6����

��
������Щ���棨���ᡢ���ᡢ���ᡢʯ���ӡ��i�ѡ��ڻ��ȣ���β�в���һ���������β��Ϊ���Բв�λ������β�����γɹ�����ǰ�������벿δ�����϶��ػ��Ľṹ��һ����β��������ѹ�����Ȼ�е�̼�ʱ���������Բв�λǰ�����β���ֱ�����ͬ������ǿ�ҵIJ�Э�����������Ǿͻ���β�ǵ�ij���Բв�λ�����ѣ���ͬ�����Ƥ��һ�����ԲУ�autotomy����β���������Բв�λ��ϸ��ʼ�ձ�������ֳ�ֻ���������˲�β��������³�������β��ͼ18-7����

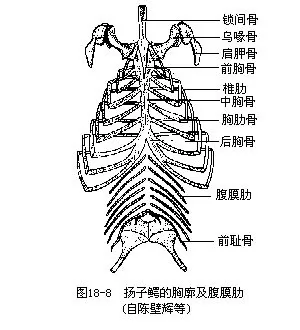
���������������������������߹ǣ�ÿ���߹�һ���ɱ��ε�Ӳ�Ǻ��ε����Ǻϳɣ��������������⣬���ж���ǰ��һ���������߹Ǿ��븹���ߵ��عǣ�sternum�����ӳ�������throax��������Ϊ��Ĥ���������У����뱣���������ٺͼ�ǿ�������õĻ���������صģ�ͬʱҲΪǰ֫�����ṩ�˸��ŵ㣻�߹Ǹ����伡�����ǵ���������������н����Ե���չ����С��Эͬ�����˶�����ɡ������Ш�������عǺ��и�����Ƥ�µĸ�Ĥ�ߣ�����Դ����Ƥ���˻��ǰ壨ͼ18-8����
����2.��֫����
������1���������һ��ʮ���ε����عǣ��������ǣ�interclavicle�����ù��ڹ������ת��Ϊ�����ڰ壨entoplastron����
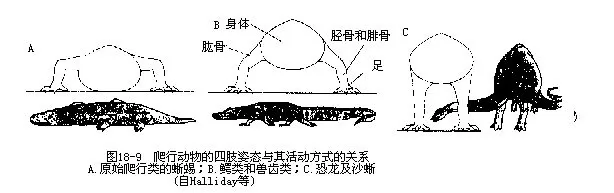
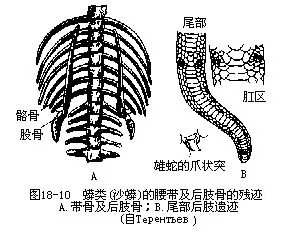
������2����֫������ij���ʺ����ֱ���ཻ��������dzС����˲������ж�����ͣϢ������ʱ���������Ÿ������ص���̬��ֻ��ɳ�����ʹ��֫�⣨elbow����ϥ��knee�������µIJ��֣�ת���·���������̧����漲�۱��ܣ�ͼ18-9������֫����ؽ�λ�������ƹ�֮�䣬���ֹؽڷ�ʽ��Ϊ�Ƽ�ؽڣ�intratarsal joint���������ࣨOphisaurus����֫��ʧ���������д��ǣ����������֫�����ǣ�Ȼ�����ơ�ä�߿ƣ�Typhlopidae������ä�߿ƣ�Leptotyphopidae����������ڣ��������˻����ĹǺɹǣ���йֳ�������һ�Խ��ʵ�צ״ͻ��Ҳ�Ǻ�֫�м���һ���֣�ͼ18-10����
�������ģ�����ϵͳ
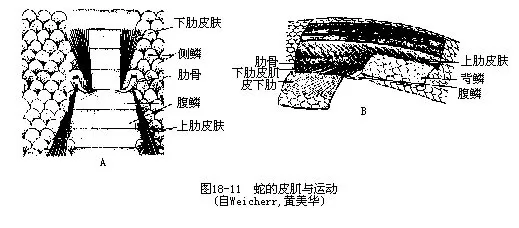
����1.Ƥ����skin muscle��һ���������ɼ�����֫�����ʲ������ֹ��Ƥ�����������ж�����Ƥ��������Ⱥ��Ƥ������������Ƥ�����丽������Ƭ��������Ǿ��нϴ�Ļ�ԣ���Զ�˺��жθ�������������Ƥ����cotocutaneous�����ֱ������Ƥ����ǰ����������������ʱ��ʹ�����˶�ƾ�巴���õ�������ʹ������������ǰ����ͼ19-11�������⣬�����ľ���Լ��Ҳ����Ƥ����
����2.ҧ��ʼ��������ֹ��λ��������temporalis����ҧ����masseter������Ϊ�տڼ�����ֹ��������Ǹ���Ķ�����Ϊ���ڼ�����ǰ���鼡���������ҧ�����ij��֣�����ʱ�����������������ڣ�ʹҧ�����ܵõ��˼�ǿ���������������ʹ������ǰ�����Ҹ���������˶�����ǿ�˲�ʳ������
��
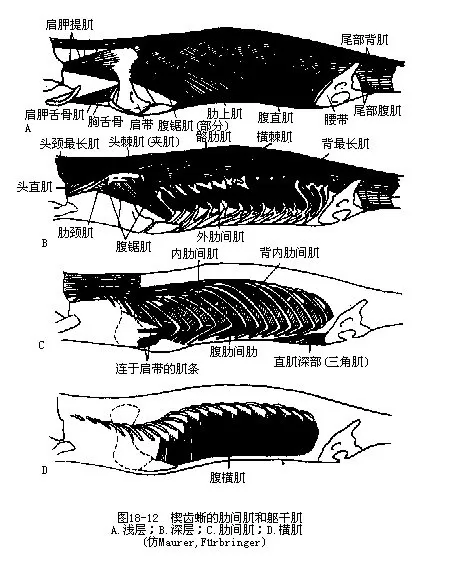
����3.�伡��intercostal muscle����Ӫ�ظ�ʽ������½�ܼ���������м��⡣�伡λ���ز��������ϼ��·���������ö�߹�֮�䣬�ɷ�Ϊ���伡�����伡�����ڵ����߹������������ظ�ǻ������仯����Эͬ���ڼ�����ɺ������ã�ͼ18-12����
����4.���ɼ� ���ɼ�����֫���������ή������������Ҫ����������������ż����ϡ��������Ļ��ܡ�����������ֻ���һ�㱡Ƭ����Ϊ������iliocostales����������չ�����ڣ�ֹ���߹Dz��档�������������������������Ե������������ͷ��������ת���йء������ɱ�������Ϊ��б������б�����ἡ���ɣ���ֱ��Ҳ�������á�
��
����5.��֫����֫�ϲ��ļ���ִ�ǰ�ۼ�������Ա�������ࡢ������米�������Ǽ�����ͷ���ȣ�������ʱ��ǰ�ٺ���չǰ֫����֫��λ������֮��ij����ɼ������ּ����Ȳ��Ĺ��ּ����β�����ȣ���Ҫ�����ǰѴ��������ڲ��ʹϥ�ؽڱպϣ���������̧����沢��ǰ������
�������壩����ϵͳ��ǻ���������Էֽ磬�з���Ĵ��٣�labialgland�������٣�palatinegland�������٣�lingual gland���������٣�sublingual gland���ȿ�ǻ�٣��������������ʪ��ʳ������ʶ�������ɡ������ʾ��������βʯ���ӣ����о������ߡ����棩��ʳ�������ۣ������á�����������������ζ̿������������еĴ���������ճ���ڿڵײ���������⡣�����������ƽ���Բ���������ԡ���������ϸ���������������ڣ�����ȱ��ζ�٣�����ζ�����ã����ܰѿ����е���������ܸ������ϣ���ͨ�����е��������֪�������������࣬��Ϊ�ݼ�����Χ����������������ճ�ԣ�ƽʱ��ѹ���ڿ��е��������ʳʱ�����ڿ��ٳ�Ѫ������ǿ������������ͷ�ӿ���ֱ���ȥ��������������Ȳ������˲�䣬���Դﵽ�������ͬ�ij��ȣ�����ȷ�����ճס���
����������������ݶ��Ե���н����⣬�������ж�������ֱ��и�����ʽ�����ݣ����������ɲ��ϸ��³�����������ͨ����������λ�ò�ͬ���ɷֶ����ݣ�����ᡢɳ�ᣩ�������ݣ���������漰�ߣ����Ͳ����ݣ������ࣩ��ͼ18-13�������ж����������Ҫ�������ϡ�������Ҳ���Գ������������Ǻ������ϣ�Ш�������������ǡ����Ƕ������ݣ��������������ܶ���һ�¶������ж�����ȴ�Ǿ����еġ�������������Ϊͬ�ͳݣ�homodont����ֻ�������������������棨ɳ�ᣩ�г����ֻ�Ϊ���ͳݵ����ơ�������fangs���Ƕ���ǰ�Ǻ������ϵ�������ö�ػ��Ĵ�����������й��������йܶ��ֱ��Ϊ������grooved tooth����������canaliculated tooth����ͨ�������������Ŷ������붾�٣�toxic gland��������������������λ�ò�ͬ����ǰ�����ͺ���֮�֡��������ϴ���ת��������һ��λ���ۺ��ڽ��Ϸ��������࣬�����⸲�м��͵Ľ����֯���ڲ��ѹ���ټ�����ѹ�����ټ�ǣ�������������á�����ҧ���ͬʱ����ͨ������Զ��ٵ�ѹ������ʹ���еĶ�Һ�����Ŷ����ܣ��������Ĺ������ų���ע�벶�������ڣ�ͼ18-14����

 ��
��
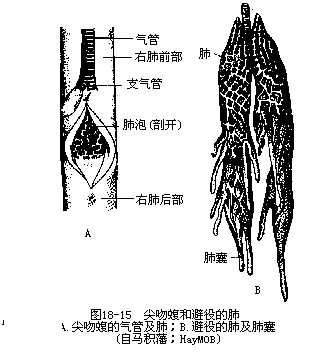
��������������ϵͳ�ڱǿ׳��ѷ�״�����������в����ʺ����������ž��ķֻ��������������˹ܱ������ǻ���֧�ֵ����ܣ�trachea�����������˷ֳ�һ��֧���ܣ�bronchi���ֱ�ͨ�����ҷ����ڡ�������һ��λ���ظ�ǻǰ������ĺ������٣�������״���ڲ����и��ӵļ����ʹ֮�ָ�����������״С�ң����ֲ��ż���ḻ�ķζ����ͷξ�����Ѫ�ܣ���������ܶ����ܸ���Ч������������Ӵ�����������ı��������������������֫���ж����У������λ�ó�����������Գ�������δ��ή�����˻����۾��߿ơ����߿ƺ�һ�������߿����ࣩ����Ϊʧȥ�������ܵIJ������١����ߺͱ�����ķ���ṹ�Ƚ����⣬ǰ��Ϊ�����������ڱ�ƽ������������������ң��ֲ�������䣬��Ϊ�������������������ã����ֽṹ�ڽ���������ʱ��ø����Ƶķ�չ��ͼ18-15����
��
�������ж�����������ܶ���ɽ����ڵ��˶����п���ʽ�����⣬ͬʱ����չ����Ĥ���ﹲ�е��ظ�ʽ�������ظ�ʽ�����������伡�����������߹ǣ���չ��ǻ������������Σ������伡����ʱ����ǣ���߹Ǻ���ǻ��С�������ӷ��ں������������þ���ͨ����ǻ�н�������ź���С�Ĺ���������彻���ġ�
��
����ˮ�ܹ�����ܽϳ�ʱ��DZ����ˮ�ף���ʱ��Ҫ���ʱں�ͻ����йֳǻ����ĸ����ף���Ƹ��ң���Ϊ�����������١����ж���ĺ�ͷ����β��������ƣ������������ĺ��Ŷ���һ����ƤճĤ���ţ������γ��������������������ڻ��ƺͱ��ۿƵ������ں������������ܲ����ߵͲ��ȵĺ�����guttural noises�����������Դ�ڻ�������“��—�”֮��Ϊ��������Ϥ��
��
�������ߣ�ѭ��ϵͳ ���ж����ѪҺΪ����ȫ˫ѭ������Ҫ�ص��ǣ�
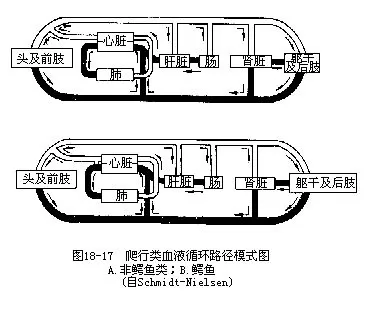
����1.����������ķ���һ���ң��������һ���ֱ��������ķ�������Բ���˻���ʧ�������ࡢ���������������Ҹ��ڳ����Ҽ����ʹ�����γɷָ�����ȫ�����ң�����������ʱ���Ҽ���ɵִ����ұ��ڣ��ڶ�ʱ����ʹ���Ҹ������������룬�������Ѫ������Ѫ�ķ���ѭ�����������������ơ���������������Ҽ�������������嶯���Ļ�������һ����ͨ2�����ҵ����Ͽף�foramen of Panizzae����ͼ18-16����

����2.���ĵ�ѪҺ�ֱ����������ҵ���������3�Զ����������зζ����������嶯�������������Ҳ�����벿�����嶯������������������������Ҽ������ȫ���γ�һ������ǻ������Ѫ�ɷξ����������ķ��������ң����в���ѪҺͨ������ǻ�����������ڣ����Դ��������в����������嶯�����ڣ�ѪҺ�еĺ��������������е�ѪҺ����һ�£���ֻ���м�����������Ѫ��ʵ���Ͻ������ҵ��Ҳ��������������Ѫ��
����3.ͨ��ͷ���ľ��ܶ��������嶯����������
����4.���嶯����Զ�����嶯�����ִ���ʼ�����ҵ����в������е�ѪҺ��Ҫ���Ծ���ǻ��ѪҺҲΪ����Ѫ����������嶯�����ϳɵı������У����Ѫ�ijɷּ��١�
����5.����ϵͳ�������ܶ�������������Ҫ��������һ��ǰ������һ���������һ�����ž�����һ�����ž������������������˻�����֫���������ѪҺֻ��һ�����ڽ�������ʱ��ɢΪëϸѪ�ܣ��������ž�������һ������Խ����ֱ�ӻ���������ͼ18-17��ͼ18-18����

�������ˣ���йϵͳ���ж�����������Ĥ����һ�����Ժ�����metanephros����Ϊ�������١���������������벿�ı��ڣ����Ҹ�һ����λ��������������в�����ȫ�Գƣ�������������ǰ������������������죬�����ʷ�Ҷ״�����ж�������Ļ����ṹ���ܣ�ͬ���ܶ��ﲢ�ޱ������𣬵��������ڵ�����λ��ĿȴԶԶ���ں��ߣ�����������Ϊ��ǿ����ͨ��ר�õ���������Һ�����侫�ܻ����ѹܿ��ڵ�ǰ�ڲ����йֳǻ��ͼ18-19����

��
�������ж��������������⣬йֳǻ�ĸ�����а��ף���Ĥ����İ�������̥�ڵ����һ��������γɣ��ʳ����Ұ��ף�allantoic bladder������������ж������ŵ���Һ�У��京��������Ҫ�����ᣨuric ac-id���������Σ����DZ����أ�urea��������ˮ�����Գ�����Һ�г����ɰ�ɫ���̬���ʡ��������ʵijɷֺܸ��ӣ������ơ����Լ�����İ��Σ����е��ơ������ӳ��������Σ�urate salt������ʽ��ͨ��йֳǻ�����ų�����ˮ������Щ���ʳ���ʱ���ֱ����ܡ��Ͱ����������ս���ѪҺ�ڣ������ٲ�����ͳ����������ظ���ת���ڸɺ�������������ж��������Һɥʧ�ͱ������ڲ����γɸ���Ѫ������ѹ��������ʮ����Ҫ����Ӧ���塣
��
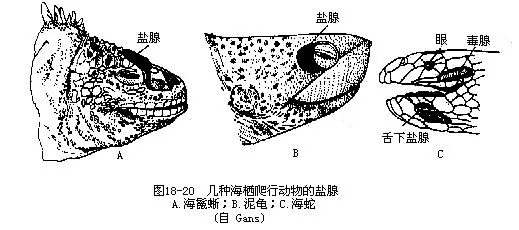
�������ꡢ�����ᣨAmblyrhnchus cristatus�������ߺ���Щ����������������Crocodylus porosus��������Crocodylus acutus���ܺ���Alligator mississippiensis���۾��� Caimen crocodilus�ȣ������������������——���٣�salt gland����ִ����������й��extra renal salt excretion�����ܣ�ͨ�����ٷ����ォѪҺ�ж�����ηִ������⡣���ٵ�λ���Ӷ����������ͬ������λ���ۺ��Ϸ���������λ�ڱDz������⣬���ڵĺ���Һ�徭С������ǵ������γ��Ȼ��ƻ��Ȼ��صĽᾧ�����ߵ�����λ�����£����������λ�����в��������࣬�����������ټ��е����٣�Ҳ�и��ӵ���ĭ״�٣�����Լ100�����������κͷ���ճҺ�Ĺ��ܣ�ͼ18-20����������Ϊ��ijЩ���ж����У����ٵ���Ҫ�������������࣬����������ˮƽ������ƽ�������Ҫ���塣
�������ж�������̥�����ڼ䣬��Ĥ�ѵĽṹ�����ܱ����������Ч����ֹ���ڵ�ʧˮ�����һ����Խ�С�������ʢ��ͨ���������ų��������εȴ�л���
 ��
��
�������ţ���ϵͳ���ж����Եĸ����Ѿ�������ȫ������ͬһƽ���ϣ����Է�չ���������������������Եس������������ԣ�Ȼ������ͼӺ����Ҫ�����Ծ����ڴ��Եײ�����״�壬����������Ҫ���Ѿ�����ͬ������Ҷ���Ტ���ˣ�������Ķ��ڼ�����������ϻ���ԭ��Ƥ�����ǿ�ʼ������ϸ����pyramidal cell�������ۼ�����ϸ���㣬���ɴ��Ա��������Ƥ��neopalli-um��������Ƥ�����ж������д���ϵͳ���������ڽΡ������ڱ��漸�����ڱ�ʶ���ɴ˷���ϸС�������٣�epiphysis���Ͷ�����parietal organ�������Ա���Ϊһ��Բ�ε���Ҷ����Ȼ�����ж���ĸ������࣬���������������Ե�����ά�У��ֶ�ֱ��ͨ����Ҷ������Ҳ����������ά�����Դ��ڴ��ԣ�ͼ18-21��������������ж������������������е����ƣ���չ����������ﵽ�˸߷塣���������ߺ���β���ѷֻ�Ϊ�ĵ��壨corpora quadrigemina����ͨ��ǰ��һ����Ҷ�������С��С�Ա����ܶ��������������������ж��ﻺ���������ϰ���йء�������Ӿ��ˮ�����ж������ࣩС�Է������ã����Եһ�㶼���������������ѵ�ǰ�벿���������С��������������ʼ�ֻ���һ��С�ε�С�Զ���������������С���ܵ�ǰ�����������ƽ����أ�ͼ18-22����
�������ж��↑ʼ����12�������������ļ�����spinal accessory nerve���Ѿ��������ֻ�������������Χ�����ڡ��ߺ������������11�ԣ��ߵ������ͼ�������δ���룬������ļ��������Ժ���������δ�ɶ�����������

������ʮ���о�����
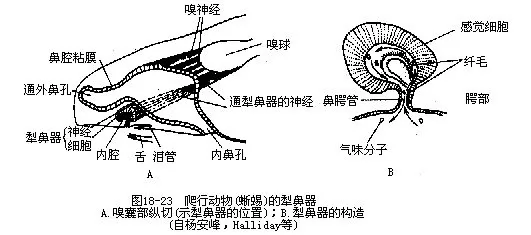
����1.��� �Ǽǣ�conchae�������ж����ǻ���״γ��ֵĽṹ�������������ѷ�չ������3���Ǽǵ�ˮƽ���Ǽǵı��渲������Ƥ���ֲ�����������о�ϸ������������������������ıǼDz����
���������������������Jacobson's organ��ʮ�ַ���Զ����ı���ܿ����ڿ�ǻ��������������ڱڸ��ио���Ƥ����ճĤ����ͨ��λ�ڸо���Ƥ������������ķ�֧������������˾���������������������ֱ����ͨ����������ⲿ�����������Ҫ����Ļ�ŵ�����ɡ�������ߵ���⾭�����ϴ����еķ�϶��Ƶ�������������ּ��֪̽���Ѽ������еĸ��ֻ�ѧ��ζ���ʣ���������ؿ�ǻʱ�����������������ѧ�����ܽ�����ճĤ�ϣ����������ͼ18-23�����������������������������������ֻͻ���ǻ����������������ͨ���ǻ��
����2.�Ӿ� �����ߺ��������е����ᡢ�ڻ��ƶ����⣬���ж��ﶼ�л�Ե��ϡ���������˲Ĥ���ڹ���ࡢ����������г��������٣�lachrymal gland����������ᆳ������ɱ�ǻ�ų������۱������һ�����ı�Ĥ�������ϡ���������չ�����Ϻ��ػ����ɣ��б�����������ã���Ƥʱ����ͬȫ���Ľ�����һ���������ɵ���
��
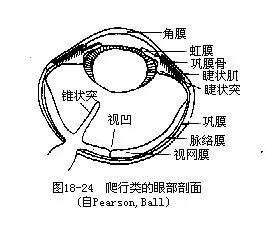
��������Ĺ���Ƚϵ��ͣ��Ӿ��ĵ����������ı侧��������Ĥ֮��ľ����Լ��ı侧�����״�����Ƶġ������Գɱ�Բ�Σ����������������Զ�ӣ�����Χ��“���”�����������Ҿ���Ľ�״�壨ciliary body�������к��Ƽ����ʵĽ�״����ciliary muscle�����������ƶ������ǰ��λ�ã������Ӿ࣬����ʱ���ܸı侧����ȣ�ʹ֮��ΪԲ����Ӧ���ӡ������ڵĽ�״���˻���ʹ������ε��Ӿ�������ã��ɺ�Ĥ��Լ����������Ϊִ�С�
�������ж�������ĺ��۷���ͨ����һ��������Ĥͻ����״ͻ��conus papillaris������Ҫ�ɽ����֯���ɣ��ھ߷ḻ��ëϸѪ�ܣ���Ӫ������Ĺ��ܡ�����Ĺ�Ĥ����һȦ�ʸ���״���еĻ���С��Ƭ��Ϊ��Ĥ�ǣ�scleral ossicle��������ijЩ�����ж���������������״��Ҳ�����ִ����������е��������б�����������ã�ͼ18-24 ����
����Ш�����һЩ������ơ�����ƣ������������Ժ�ͷ�����У��е������ۣ�parietal eye��������λ�ڼ��Զ����Ķ��ˣ�����Ƥ�£�ͨ���������ϵ�������Լ�ɼ�������Ҳ���������롣���۵Ļ����ṹ�����������ƣ����к�С�Ľ�Ĥ��ˮ���������Ĥ�����Dz���ʹ����������Ĥ�ϳ����ий�ܡ����۶���������ں��ʶȵ������չ��������ż�����Ҫ�����ã���������ijЩ�����Ե��������ʶ���;�������й�ϵ�����ж���Ķ�����Դ�����ɹ��壨parapinealbody���������������Ķ���ȴ�����ɹ��壨pineal body=������epiphysis����������Ϊ��ԭʼ�ļ�������ܴ���2�����ۣ��������з�ʽ����һǰһ�������ҶԳƣ���Щ��Ƥ����ͷ�ǻ�ʯ�Ľṹ�����Գ���֤ʵ��һ���۵���ȷ��
 ��
��
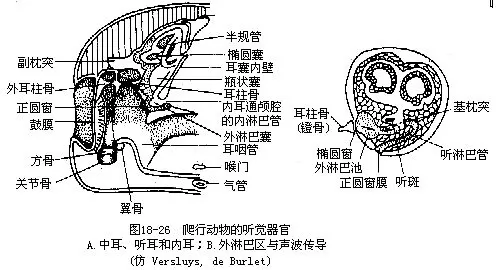
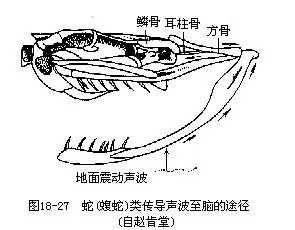
����3.���� ���ж���������������ڹ�Ĥ���ݶ������˳��ε���������������ж����ж����ǺͶ��ʹܣ������ǵ��⡢�����˷ֱ�Ӵ���Ĥ�ͽ����ڶ�����Բ����fenestra ovalis������Բ��֮�»�������ǰ��δ�е���Բ����fenestra rotunda����������б�Ĥ��ʹ�ڶ����ܰ�Һ���������˻�����ء��ڶ���Ĥ��·���������ܶ��������ͬ������˾������ƿ״��������������ƿ״��Ļ�����һ���ɸо�ëϸ����ɡ��������ʻ�Ĥ�ڸǵĻ���ͻ��Ϊ���ж������Ҫ������������ͨ����Ĥ�����ǵ�������Բ�������ڶ�������һ�����ܰ��ƹ�ƿ״��Ļ���ͻ�ִ���Բ�������ܰ������ͻ�µĸо�ëϸ������һ����Ļ�Ĥ�������Ĥ���ɼ�����о�ëϸ����ͬ��������ëϸ���·����������ڸ�֪�����������ƿ״���ӳ��ɾ����Ķ��ϣ�cochlea���������������������������������������ڷ�ֳ���֮���Ժ�������ѶϢ��Ҳ��֤�����������ٷ��ͼ18-26����������ӦѨ������ж�����Ĥ�Ͷ��ʹܾ����˻������Ƕ������Ա�������������������ؽ��ܴӵ�����������������ͨ�����ϵķ��Ǽ���֮�����Ķ����Ǵ����ڶ����Ӷ�����������ͼ18-27����
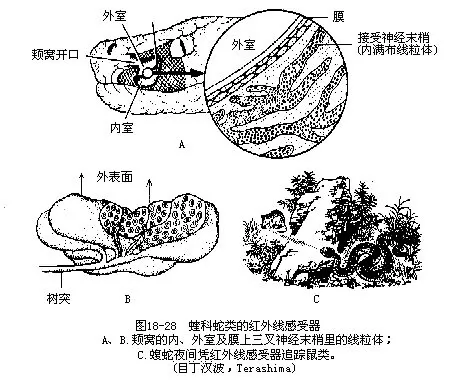
����4.�����߸�������infrared receptor������������ǿƺ���������ͷ�����е����ܸ���������λ�����ߡ���Ҷ�ࡢ��β�ߵ��۱�֮��ļ��ѣ�facial pit�������ഽ�۱���Ĵ��ѣ�labial pit����������ļ�����һ�������ΰ��ݣ�������һ����10μm��15μm�ı�Ĥ�����ѷָ�Ϊ�ڡ������ң����ҽϴ��ڳ��⣻����λ�ڼ��������һϸ�ܵ�����ǰ�ǣ���һС��ͨ��Ƥ�����棬�ɵ����ڡ�����֮���ѹ����������Լ��������Ĥ���ڱ����ܲ���������֧����ĩ�ң����ն���Ϊ����ڲ����������壨mitochondrion��������һ�ּ������е����ܼ������ֻ��123.68×10-5J/cm2������������ʹ֮�����֪��һ����Χ�ڵ������������35ms��˲Ϣ֮��Ѹ��������Ӧ����Ч��Ϊ�ֽ��������ĺ�����̽���������������ǿ����ڹ۲����Ĥ�ϵ���ĩ��ʱ�����������������̬�����������͵���Ӧ�ı䣬��ͱ������ܸ�������������������Ĺ�ϵ��Ϊ���С��ߵļ��Ѳ����ܾ��쵽��Χ�����е���Ѫ�����������ȷ���ö����λ�ã��Ӷ������ٺ���Ϯ�������Ҳ����Ϊһ��������ҹ����ʳ������о����٣�ͼ18-28����������λ���������ۻ��ϴ��۱����С�����ݡ���ṹ�����ǿ�����ļ������ƣ�Ҳ��һ���Ȳ�λ�����ܸ�֪0.026����²�仯����������ѷ�ڼ��ѡ�
��
������ʮһ����ֳϵͳ ���ж���Ӫ�����ܾ�����������������Ш�����⣬���Ծ��н��������ߺ���������Խ�������һ��������hemipenis����ƽʱ����¶���⣬�����йֳǻ��λ��β�������2�����ʵ��������У�����Щ���������β�������ԵñȽ�����������ɱ���Ļ������ɣ�����ʱ��ѹ����������ʹ֮����йֳǻ�⣬�ҵ�ĩ�˽��ʹ�������β���ϡ��������ĸ�����������ľ�����������ǰ���ֲ������˵Ĺ�ͷ������ʱ��2��������ͬʱ��йֳǻ�ڷ���������ֻ��һ��İ������������йֳǻ�У��������Ļ�����йֳǻ��ͨ���ɽ�˳�侫�ܶ��µľ�Һ�����Ű������ľ�����ע��������ڡ����ֶ���İ�������̬��һ������Ϊ���༰̽���������Ե��ϵ�����ݣ�ͼ18-29����
��
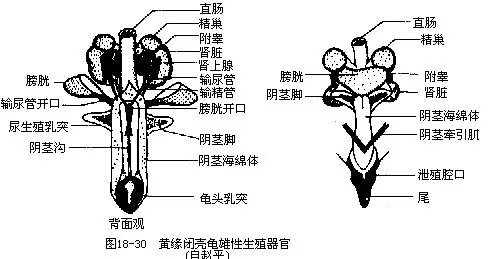
�������Թꡢ���йֳǻ�����е�������������Ϊ������penis�������к����壬�ܳ�Ѫ����������⽻�䣨ͼ18-30����
��
�����������ѳ������ѹܸ�һ�ԡ����ѹ��϶�Ϊ��������ǻ�����ȿڣ��ж����е����ٶ���Ϊ�����ڲ�����������������٣����¶����ܷ����γɸ��ʣ���������ࣩ��ʯ���ʣ��ꡢ������ѿǵĿ��ٲ������ѹ�ĩ�˿�����йֳǻ�����ԵĹ�ͱ���йֳǻ������һ���������Ե����٣�clitoris����������������ͬԴ���ٵĽṹ��
��������������ж�����������ʽ��ֳ����Ҫ����������¶Ȼ�ֲ�︯�ܷ��Ͳ������������з�����
������Щ�ű����γ�ȸߺ��ε������ߺ����棬�Լ�������Ӫ��̥����ovoviviparity�����ܾ��������ѹ��ڷ���������ʱ�������������ࣨʯ���� Eumeces chinensis����βʯ����Eumeces ele-gans�����ܾ�����ĸ������ѹ����ѳ���������������ǰ�ɽ��������γɽΣ������������ݼ��۵�ȣ��ǽ�����������̥��֮���һ�ֹ������ͣ��ɳ�Ϊ����̥����������֪���ҹ�������12�������25����Ӫ̥������̥����Լռ����������7.8����ռȫ�����ֵ�13.7���������������Ͻ�ɳ�ᡢ�i�ѣ�Lygosoma indica�������뻬�ᡢ�ܵ����ᡢ���ᡢ��ä�ߡ�ɳ���������ߡ�ˮ�����ߡ�ˮ�ߡ���ԭ�����ߡ��״���Ҷ�ࡢ���ߡ�Сͷ���ߺͺ����ߵȡ�
��
�ڶ��� ���иٷ���
��
�����ִ�����ж���Լ��6550���֣��ҹ�Լ��380���֣����ͷ��Ŀ�⣬����4Ŀ���ھ��зֲ���
��
һ���ͷ��Ŀ��Rhynchocephaliformes��
��
������Ŀ�����ж���������ϵ���Ⱥ֮һ�����Ǵ���������¶����ͺ������ͣ������㷺�ֲ���ȫ���ϱ���ŷ���ǡ��Ǻ��������ޣ�����ֻ��Ш���ᣨSphenodon punctatum��1�֣����������������¿˺�Ͽ��һЩС�����ϣ��ܹ�����ǧβ���ѱ��پ���ı�Ե������Ш�������һϵ�����ƹ����ж����ԭʼ����������ڶ���ѧ����“�ʯ”֮�ơ�
����Ш�����������ᣬ����ͷ��ǰ�˳����״�������ݣ��������ͷ�ᡣ�峤50cm��70cm���屻С�����ۣ�ͷ�����ͱ��������һ���о��״���ۡ���������ɫ������а�ɫ�ߵ㡣Ш�����ԭʼ�����У�˫�����ǣ���������м����IJ��ࣻ�����ļ������и�Ĥ�ߣ�������������ͷ��Ϊ˫���ͣ����������������ϡ��������Dz��ܻ�������ݣ�����ʮ�ַ�����о��塢��Ĥ������Ĥ�������۵Ľṹ��
����Ш�����뺣����“���Ṳ��”�����������ҹ���������ס�ں��ס������̡����ࡢ����˵Ķ�Ѩ�ڣ�ҹ�������ʳ���桢������Ϻ�ȣ�ͼ18-31����11��������2�¹�����8��15ö��������Լ13���¡�������������������20�����Ҳ��Գ��죬��������77��ļ�¼���ݳ�������ɴ���ꡣ

 ��
��
��
�������Ŀ��Testudoformes��Chelonia��
��
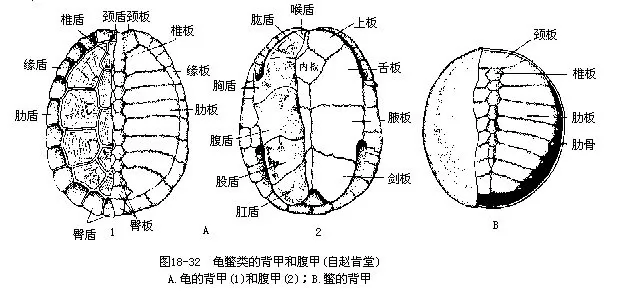
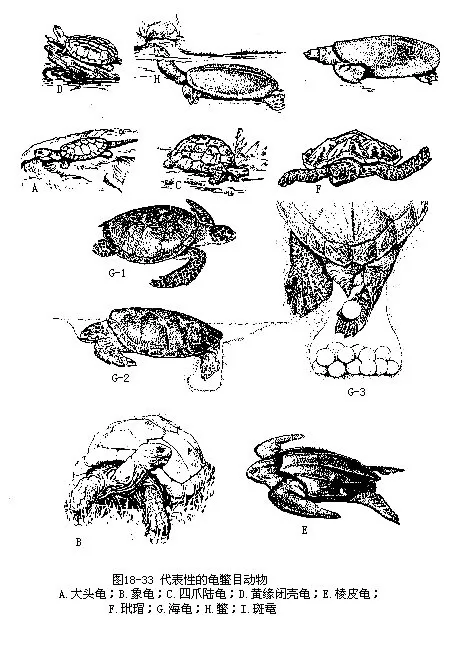
�������Ŀ�����ж����е��ػ���Ⱥ��������̣����ɲ��������ڼ�̵Ĺ���Ӳ���ڣ�ͷ��������֫��β��¶��������������������С�Ӳ���ɱ�������ɣ��⸲���ʰ����Ƥ��������������ͬ�߹�һ���뱳�������ϣ����λ���߹ǵĸ��棬���Ǽ������о����е��������عǣ�Ҳ���γ����������������عǺ����Ƿֱ�����˸��ף��ڰ���ϰ壩����ɡ�����û���ף����������Žᣬ���ܻ���Ϳ����ݣ���֮���Ե�Ľ����ʣ����������ԣ���˲Ĥ�ͻ��������йֳ�����ѻ�Բ�Ρ����Ծߵ������������ѿǸ��ʻ���ʣ����꣩��ÿ�����2��3�Σ�һ�εIJ�����Ϊ��ʮ����200ö���ϡ����������ϳ���һ��ɻ���ʮ�꣬�ݱ����꣨Testudogigantea���ͺ����������ɴ�150��250�ꡣ�ִ�������250���֣�������13�ƣ����ֲ����´����ȴ��������ҹ���37�֡������Ե������У�ͼ18-33����
������һ��ƽ�ع�ƣ�Platysternidae�����ƽ�1��1�֣���ƽ�ع꣨Platysternon megacephalum����ͷ���ǿ���������ӥ��״���׳ƴ�ͷ���ӥ��ꣻβ����Լ�븹�׳���ȣ�ͷ��֫��β�����������ڣ���DZ�ƽ�������������ʹ����ӡ�������ɽ��Ϫ���У�ҹ������ƾ�平���צ�ͳ�β�İ�����Խ���ϰ������Ԯ�������ֲ��ڶ����ǵ������ҹ�����������ʡ��
������������ƣ�Testudinidae��ͷ��������ͷ��ǰ�����ۣ�������а��ݡ���Ǽ�Ӳ�������������ı������ڼ��Ŵ��ԹǷ�Ӻϣ����տǹ꣨Cistoclemmys������꣨Cyclemys������ʹ���������ǵĹǰ��⸲�Ա�Ƥ�ԽǶܡ�ͷ��������֫��β����������ڡ���֫��׳��צ�۶�ǿ��ָ��ֺ����������룬Լ��90���֣����������⣬�㷺�ֲ���������ء����д����Ե������У�
������꣨Geochelone elephantopus������¡���ߴ�50cm���ϣ�ȫ��140cm������Լ 250��kg��������������½�����ࡣ�ֲ�����̫ƽ���϶����¡Ⱥ����
������צ½�꣨Testudo horsfieldi��ͷ���жԳƴ��ۣ��Ƕ̣����װ����ζ��ߣ���֫Բ��״��4צ��ָ��ֺ�����룻β�̣�ĩ����һ���ʵ�צ״��ڡ������ҹ��½����ǵ�����ɳĮ�У�ÿ��4��7��Ϊ����ڣ��ڶ����ӣ�����������������¾�����ɳѨ����������״̬���к���֮�ơ�
��
�����ڹ꣨Chinemys reevesii���������Ͳݹ꣬���ҹ�����Ĺ��࣬��������������ظ�ԭ�⣬������ʡ���зֲ���ͷ��ǰ��ƽ���������ۣ��Ե�Ľ��ʽ�����������3�����⣻ָ��ֺ��ȫ�롣�����ڽ��ӡ����Ӻͳ����ڣ�����������İ����������ڶ����ѡ���ʳ�ԡ�
������Ե�տǹ꣨Cistoclemmys flavomarginata��ͷ��Ƥ���⻬����ĤԲ����������״�����ļ������ԣ������ضܺ���������Ӧ�Ĺǰ�֮�����ʹ�������ʹ����ǰ����������Ի����ͷ��β����֫�������ʱ����ȫ�պ��ڱ��ף��տǹ�����Ƽ��ɴ˶�����ָ��ֺ�����룬�߶�צ������������Ե��Ե�ܸ��������ɫ������ˮ���������ҹ����½�����࣬��Ҫ�ֲ��ڳ������ϵ�����
�����ƺ�ˮ�꣨Mauremys multica��ͷС�������⻬���ۣ��Ƕ˳���������ƽ̹����Ե��һ���������ָ��ֺ��ȫ�롣β��ϸ���ʲ����ɫ��ͷ�����ۺ��ع�Ĥ�ϡ��¸���һ����ɫ���ơ������ҹ��Ϸ�����ش���ɽ����ˮ���У����ڸ�����С��ľ���ݴ��ڻ�����䱳���н��ʣ����Գ��и�ë��ȸ�����֮��������Ϊ“��ë��”����䲶�˹������͡�
��������������ƣ�Chelonidae�����ױ�ƽ���Գ������Σ��߹dz���ĩ��������Ե���������������С����������֮����ʹ�������ͷ��������֫����������ڡ���֫��״��ָ��ֺ�DZ�ƽ��������1��2צ��������ůˮ�Ժ������㡢Ϻ��ͷ��١��Ƕ���ͺ���Ϊʳ����ֳ�����ϰ�����Ͳ��ѣ�ÿ�β��Ѱ���ö���ѿǸ��ʣ�������40��60�졣���ƹ���6�֣��ҹ���4�֣��������ǣ�
�������꣨Chelonia mydas���峤Լ1m����Լ300kg��400kg������Ƭƽ�������ǰ�˲��ɹ���������Ե�о��״ȱ�̡�ǰ֫�����ڲ���һצ��β�̡���ֳ���ڵĺ��꣬ÿ�궨�ڳ�Ⱥ����Ǩ��ңԶ�ĺ����������䡣�ƹ���ҹ�����ϰ���ɳ̲��Ӳ��ѣ�������ɳ���Ǻع麣���ѽ�������������з�����ÿ����Ѷ�Σ�һ�β���90��160ö��������̫ƽ��ĺ������������������ҹ���ɳȺ�����з�ֳ���ѡ�
������裣�Eremochelys imbricata���Dz�⣬������ӥ�졣�걳�����гɸ���״����ͬ������������ƽ��״��ǰ֫��2צ���Ժ�����ͺ���Ϊʳ�������������й��ɼ����ۻ����ӹ��ɸ��ֹ���Ʒ���ֲ����ҹ��Ϻ��Ͷ�����
�������ģ���Ƥ��ƣ�Dermochelyidae��������ȿɳ���2m����800kg�����ִ�������ࡣ�����ɼ���ö����ε�С�ǰ���Ƕ���ɣ��⸲����Ƥ�������ʶ�Ƭ����������7�����⣬ȫ������������ϳ�һ�����ε�ĩ�ˡ���֫��״����צ����1��1�֣�����Ƥ�꣨Dermochelys cori-acea���������ȴ������ȴ������ҹ��������Ϻ����зֲ���
�������壩��ƣ�Trionychidae���С�С�͵�ˮ���ࡣ���������ʣ���Ե��ͽ��ʶ�Ƭ�����������ĸ���Ƥ�������ױ�ԵΪ��ʵ�Ľ����֯���׳�ȹ�ߡ��Ƕ˼���ɿɶ�����ͻ���ǿ����ڴˡ���֫����������ڣ�ָ��ֺ�������ڲ�3ָ��ֺ��צ������Լ��20���֣���Ҫ�ֲ��ڷ��ޡ������Ǻͱ������ҹ���4��5�֡�
�����Pelodiscus sinensis��������������㡣��ͻ����Լ���۾���ȡ����С���������7kg��8kg���ʲ���ͻ������Ѫ�ܣ���������ˮ�н�������ĸ����������٣���˱�����ˮ��DZ������10Сʱ֮�䣬���ص�ˮ����������������������⣬�ֲ��鼰ȫ����ʡ��
����������Pelochelys maculatus�����ͱ��࣬���ؿɴ�109kg����ͻ�̣��dz�С���۾���һ�롣�����ڽ��Ӻͺ����У���ʳ�����ڳ������εĽ����ϲ����㽭���������ϵĶ��ϲ���Ϊ���پ������ϡ���
��
��������Ŀ��Lacertiformes��
��
��������Ŀ�����иٶ�������������һ����Ⱥ������������֫���ָ��ֺ5ö��ĩ����צ���������к��ھ�����������֫�˻���ȱʧ���м�����عǡ������ɶ������ƽ�Σ����������������ʡ����������ԹǷ����ӣ����������ݻ�����ݡ���Ĥ�����Ҽ����ʹ�һ������ڡ�½�ܣ�Ҳ�����ܡ���ˮ�ܻ�Ѩ�����ࡣ��ֳ���ڣ����᳣��Ϯ����ʽ�����ԣ�ǿ�н��䣨ͼ18-34�������ϼ����⣬�㲼��ȫ���ִ������Ŀ����Լ3750�֣�������16�ƣ��ҹ���֪Լ��150���֡������������У�ͼ18-35����
 ��
��
������һ���ڻ��ƣ�Gekkonidae��Ƥ���������屻���ۡ��۴�ͫ�׳���ֱ���������������Ԯ���������ָ࣬��ֺĩ�˾���������״��ָ��ֺ�棬��ı�����1��2�к������е�ֺ�°꣬���ϸ����ŷ���Ĵ�ֱ��ϸ˿��������������Ԯ��ĽӴ��档ϸ˿�����з��������������Ԯ��ĸ���������������ƽ������ʯ��ǽ�������С���˫���͡�β���Բ�����������ǿ�������������ҹ�����ܷ�������������Լ600���֣��ֲ��ڸ������ȴ����´��������ҹ���30�֣����������У�
��������ڻ���Gakko japonica�������ڻ����ع����dz������ϵ���ϰ����һ�ֱڻ�����һָ��ֺ��צ��ָ��ֺ�°굥�У�ָ��ֺ�����롣�屳������С���ܼ��������ڻ����Ƶ�����зֲ��ڻ�������������ڻ���Gekko swinhonis�������������DZ����������Ĵ����ۡ�
������ڻ���Gekko gekko���峤20cm���ϡ�ͷ����ı��汻���ۣ����������г��еĴ����ۡ���ڻ�������ҩ���Ǵ�ͳ��������ҩ�ģ�������ʱ�ܷ���“��—�”�������������ֲ��ڹ������㶫��������̨������ϵ�ʡ��
��������ɳ����Teratoscincus przewalskii��ͷ���۾��ղ�ͻ����ͷ��Ϊ���ۣ����Ϻ���֫��������С�ۣ�β���и���״���ۡ�ָ��ֺĩ�˾�צ������������״��������Ƭ����γ���״Ե����ư�ɫ��������β�����������غ�ɫ���ơ����ڻ�Įɳ�ػ���̲���ֲ������ɹš�������½���������������ҩ�����档
��������������ƣ�Agamidae����С�����档ͷ���Գƴ��ۣ����۸���״���У�������������ۡ������ݣ������ηֻ����ơ���ǰ���͡�β�������Բ���������������̥��������Լ��300�֣���Ҫ�ֲ������ޡ��ҹ���40���֣����������У�
�����߷��ᣨDraco maculatus�����Բ�⡣�������״ƤĤ��Ϊ5���ӳ����߹���֧�֣��ܻ�������֦�䣬��ʳ���档�ʺ��·���ɫ���ҡ��ж����ݣ��ƶ������ɶ���һ�������ϲ���2��5ö���ֲ��ڹ������㶫�����Ϻͺ��ϵ���
����ϲɽ���ᣨAgama himalayana���峤Լ30cm��������������Ĥ�·����ۣ������������ۣ�����״���У�����ƽ�������С��������ɫ��������β�лҰ�ɫ���Ƽ�����Բ�ߡ��ֲ������غ��½��������ں���2300m��4100m��ɽ���ҷ켰��ʯ�䣬Ҳ�ɳ����ں���1400m�Ļ�Į��ԭ�ϡ���������ɽ�����ڶ����ӣ��ڶ����ܾ���Ѹ�ٴܻض��ڻ����ʯ���С���ʱҲ������Ѩ�ӣ�����������ȥ���������档
������ԭɳ�ᣨRhrynocephalus frontalis��ͷ�����ϡ������۵ı�Ե����Σ�����ʱ����ֹɳʯ�˼��۾��������ۡ�ָ��ֺ�����з������״Ե����������֫�����Ӵ��������������������ɳ�ڡ��н�β˦�����������¾�����ϰ�ԡ�����ʱ�������۶��ӣ�����ɳ���Q��ҡ�����ӣ�ʹ�����³���ͬʱ�ú�֫�����ھ�ϸɳ��������֮ɳ�����屳���������С�������7�·������ᣬ�ֲ������ɹš��ӱ������������ࡢ���ĺͱ���������
����������ʯ���ӿƣ�Scincidae��ͷ���жԳ����еĴ���Ƭ��ȫ��������״Բ�ۣ��������¾�����Ƥ�Թǰ塣�����ݣ������״��β��Բ�����Բ���������������̥��������Լ��600���֣��㷺�ֲ��ڸ����ޡ��ҹ���31�֣������������У�
������βʯ���ӣ�Eumeces elegans��������ϸ����֫�϶̡�����Բ���⻬���������ɫ���� 5����ɫ���ƣ�ֱ��β����β�ĺ�벿��ɫ�����ڵ�ɽɽ�ּ�ɽ����Ե�ʯ���£�����̥����ֳ�������ҹ��������ϸ�ʡ��
�����i�ѣ�Lygosoma indicum���峤Լ90mm������β��������ʹ�ͭɫ���׳�ͭʯ���ӣ�������һ����Ŀ�ĺ�ɫ���ƣ�ֹ��β����������ƽԭ���ɽ��������̥����ÿ�β���6��8β���ֲ��ں��ϡ�����������ͳ������ϸ�ʡ��
�������ģ�����ƣ�Lacertidae��ͷ���жԳƵĴ���Ƭ�����۷��λ���Σ��ݺ����г��С���֫����й��ѻ������ѡ�β���ϣ�Ҳ�������������������ݴԡ���ԭ����Į��ƽԭ�ش�����������̥����ȫ����Լ��140���֣��㷺�ֲ���ŷ���ǡ������ޣ��ҹ���20�֣������������У�
���������ᣨTakydromus septentrionalis�������β����ϸ����ͷ���Ǽ⣬�����ף��������ԣ���Ĥ��¶����֫��ָ��ֺҲ���������ڲ������һ�������ѡ����������汻�������ۣ�ƽ�����С�������ɫ������ɫ���ֲ����ҹ����������ϸ�ʡ��
�����������ᣨEremias argus����϶̴֣���֫��С��β����ͷ�峤������ȡ����ۿ���״�����۷��Σ����⣬�������ų�б�С��ʲ���һ�����ε���Χ�������ڲ����һ�й����ѡ����������������еİ�ɫ�۰����γ��������ڳ��й�ľ�Ի�ܸܸ�ݶ���Χ��ɳ���ϲ�ʳ���档�ֲ����ҹ��������������������������½����ຣ����ʡ��
�������壩����ƣ�Anguidae��������Σ��к�֫�ǵIJ��ࡣȫ��������״Բ�ۣ���������Ƥ�Թǰ塣��С���л����������а�����ݹ���β���ϣ��������������ݣ���״��һ��������50�����ң��ҹ���3�֣�������Ǵ����ᣨOphisaurus harti���������ڲݴ��У�Ѩ�ӣ�ҹ�����������������ţ�����嶯��Ϊʳ���ж���ʽ���ߣ��������������������������ϵ�ϰ�ԡ��ֲ��ڳ������ϸ�ʡ��
��������������ƣ�Shinisuridae���������������ɴ�׳����֫���ָ��ֺĩ�˾���������צ��β������⡣ͷ����Ƭ��״��һ�����¶Գơ����������ۼ����д������ۣ����γ���������������β����2�����ա���Ĥ���ԡ���̣�ǰ�˷ֲ档�����ݡ���̥�������ƽ�һ�֣��������ҹ�������ɽ�����ᣨShinisaurus crocodilurus�����Ǽ����������ж��Ϊ����һ���ص㱣�����
�������ߣ�����ƣ�Varanidae�����;�������Ŀ����������Ⱥ�����弰��֫������׳��β������⡣ͷ���ԳƵĴ���Ƭ�����汻��״���ۣ����۷��Σ���������Ƥ�Թǰ塣��ϸ�����ֲ棬��������������ڡ������ݡ�β���Բ����������ƽ�1��30�����ң��ֲ��ڷ��ޡ������ޡ������ϲ������ϲ����ҹ���Բ�Ǿ��ᣨVaranus salvator����һ�㶼��ˮ�����½�ܣ�����ˮ����Ӿ��Ҳ����Ԯ������ʳ����ʳ�ԡ������꼾��ʼ���ѣ��Ѳ��ڰ��ߵ���Ѩ�ڻ������У�Լ15��30ö���ֲ������ϡ��㶫�������ͺ���ʡ��
�������ˣ����ۿƣ�Chamaeleontidae���������棬������������������������ٵĹ��켫���ػ������ƹ�3����85�֣�������2�ֲַ����������ϲ���ӡ�Ⱥ�˹�������⣬��������ȫ���ڷ��������˹�ӵ������ƵĴ����DZ��ۣ�Chamaleon sp.���������⣬�������ۣ������м��⡣��֫����ǰ����ŵ�5öָ��ֺ�ֳɻ�����������飺�ڲ�3ָ���ϳ�һ�飬�����ָ���ϳ���һ�飻��һ����ֺ�γ�һ�飬����3ֺ�γ�һ�飬����ץ����֦��β�������ڲ��ơ��۴�������������۶��ܶ���������������ת����������ʳĿ�꣬��Ұ��������������������һ���ĶԹ���ڣ�ֱ��˫��ͬʱ�����塣���ĩ�������ճҺ������ʱ��Ѹ�������Ͷճ�����Ƥ����ɫ��ϸ���������ʽᾧ���۹��Լ�ǿ����������֧���£��ܿ��ٸı���ɫ��Ӧ��Χ������“��ɫ��”���������ɴ˶�������������̥�����Ϸǵ�٪����ۣ���
�������ţ�����ƣ�Helodermatidae��Ψһ�ж������棬��������������������ݵ�ǰ���������������´����ػ��ɶ��١����η��֣�β�̶��֣���������״С����Ƥ���б�ƽ�Ĺ��ۡ���ɫ��Ŀ�ɲ��������Ұ�ɫ���ɫ�����а߲�����ķۺ�ɫ����ɫ����ɫ�İߵ㣬β�߿�������ɫ���ơ�����ɳ�أ��ж��ٻ��������ࡢ���Ѽ�С��Ϊʳ�����������ƽ�1��2�֣�����β���ᣨHelo-derma suspectum�����鱳���ᣨHeloderma horridum����
��
�ġ���Ŀ��Serpentiformes��
��
������ϸ������ͷ�����ɺ�β�������������ԡ���֫��ʧ�����Ǽ��ع��˻��������ơ�ä�߿ơ���ä�߿ƺ�Ͳ�߿ƣ�Aniliidae��������й��輰�����֫�����������˲Ĥ�����١�ȱ����Ĥ������ή�������ʹ�Ҳ��ʧ���ڶ�����Բ���ͷ���֮���ɶ�������ӡ������ڴ���Ĥ���˻�����ʧ������˫�ĺۼ�����ǰ���͡������β�⣬�������϶����пɶ����߹ǣ���������ǰ��������Ҫ֧�����١��ɶԵ����������������ε�Ӱ�죬ʹ�����ҶԳƵ�λ�ñ任��ǰ����λ�ã���ֻ����һ�����٣���һ����ή�����˻����ް��ס�������һ�Խ���������������̥��������ǰ����ż�Ļ�����Ϊ��ͼ18-36����
���������������ڽ��������и߶��ػ���һ����֧������ϵ���������������Ե��ϵ�Ƚ����У������ߵ���Щ����Ҳͬ�����������ijЩ�����У����������û����֫��������ͣ�����Ƶ����ֲ����һ��������ƵĶ��ײ����������Ƶȡ�ȫ�����ִ�����Լ��3200�֣�������13�ƣ��㲼�ڸ����ޡ��ҹ���210���֣������ж���50�֡��������������¿ƣ�ͼ18-37����
������һ��ä�߿ƣ�Typhlopidae������һЩ��������С�������ࡣ����������״��ͬ��Բ�ۣ��⻬���⡣�����˻���������Ƭ֮�£��ʳ�ä�ߡ��Dz��ܻ����������������ǰ�����ϣ������������ſڽǶȡ�������������ݣ�����ݡ��������ǵIJм��������������ڽ�ԭʼ�����ࡣ��������̥�����ҹ���3�֣�������ǹ�ä�ߣ�Ramphotyphlops braminus������Լ15cm�����ҹ���������С��һ�֡�ӪѨ������������ϡ����ѡ����漰���׳档�ֲ��ڳ������ϸ�ʡ��
�������������߿ƣ�Boidae������С���⻬�����۴���������������˻����������йɹDz��ࡣйֳ��������һ�Խ���צ״��гɶԵķΡ���������̥����ɳ�������е�����������з�����Ϊ��ĸ���輡��������������������£��������ѵķ�������Ҫ�Ժ��¶���Ϊʳ����������չ��������ʳ������Ӧ�����ܸ�����������Լ��60���֣��ֲ����ȴ����´��������ҹ��Ϸ������ߣ�Python molurus����������ɳ����Eryx miliaris��Ϊ���ƵĴ�����
��
�������������߿ƣ�Colubridae����������Ŀ�а�������������Ⱥ���ִ�����������2/3��������һ�ơ�ͷ���жԳƴ���Ƭ�����ۿ����������ݣ�������������Ǻ����2��4ö�ϴ�Ĺ�������������̥�����ֲ�����ȫ���ҹ���140���֣��������У�
�����Ƽ����ߣ�Coluber spinalis��ȫ��Լ0.5m���������ɫ���Զ�ؼ�����β����һ�����Ե�dz��ɫ�ݴ����ǻƺ��Ա�������������֮һ�����Ͽɴﳤ�����θ��أ���������������Ϊʳ��
���������ߣ�Dinodom rufozonatum���dz��������ߡ������ɫ����70������ɫ���ƣ�ͷ����Ƭ��ɫ����Ե糺죬�������һ�����κ��ƣ�����dz���ްߡ����㡢�ܡ���ܡ����桢�����С��Ϊʳ���ֲ��ڻ����������ͳ������ϸ��ء�
������ü���ߣ�Elaphe taeniurus��ȫ����1.5m���ϡ��ۺ���һ����������״���ü��������������ũ�ἰ�丽�����ء��������Խ��ױ����ܾ�ʱ������ͷ����������������ʳ���࣬�����������ߡ��ֲ��ڶ����������ͳ������ϸ�ʡ��
�����������ߣ�Rhabdophis tigrina�����氵��ɫ������ǰ�������н������ĺ�ɫ�ͽۺ�ɫ�߿飬�������ɫ�����ˣ��������ɫ��������ˮ�߲ݴԣ���ʳ�ܡ������ࡣ
���������ߣ�Zoacys dhumnades���������ߺ��ڷ��ߡ����ĸ���ȫ���ɴ�2m���ϡ������̺�ɫ��ں�ɫ������2��4�б������⣬������2���ݹ�ȫ���ĺ��ƣ�����������ں�����������ԡ�����������ش�����Ұ�䣬��Ҫ����Ϊʳ��Ҳ��������档�ֲ��ڳ������ϵ��������ҹ��Ĵ�ͳ��ҩ�ġ�
�����й�ˮ�ߣ�Enhydris chinensis���������ߡ�����Բ���Σ�ͷ���Ա�����ھ���������ƽ����β�̣�β����˫�С��ǿ׳��ϣ����ڰ�ˮ��������2ö����Ϊ�ϴ�Ĺ������������ɫ�����ڶ���������Ƭ��ɫ���γ����������ڵ��������ˮ���ȴ�����ʳ���ࡣ�ֲ��ڳ������ϵ������DZ����е��ж����ࡣ

��
�������ģ��۾��߿ƣ�Elapidae����������һ�����߲����������ǵ�ǰ����һ�Խϴ��ǰ������������м�öԤ�����������Ƶ��߶���Ҫ�������˺Ͷ������ϵͳ�������ࡣȫ����Ķ����У���һ�����ҵ���������ڱ��ƣ�Լ180���֣��ֲ��ڴ����ޡ����ޡ��������ޡ��ҹ���9�֣�ȫ�ֲ��ڳ������ϵ��������������У�
�����۾��ߣ�Naja naja��ȫ��1m���࣬�����ں�ɫ������խ�Ļư�ɫ��ߡ���ŭʱ������ǰ���ܰ���ֱ��������������ƵƵ��ǰ�ڶ�������״���������ʹ�ô��İ�ɫ���Ƴ����۾�״������������ƽԭ��ɽ��ɭ���У�ҹ������ʳ�㡢���ܶ���ͳ���ȡ��ֲ��ڻ��ϸ�ʡ�������͵Ķ���֮һ��Ҳ���������˵���Ҫ���֡����۾��߽��ƵĻ����۾����ߣ�Ophiophagus hannah����ȫ��2m��6m����Ҫ��������һ�Դ����ۡ�
���������ߣ�Bungarus multicinctus��ȫ��1m���ҡ�ͷ����Բ�Σ��Դ��ھ���β�Ҽ�ϸ������ߺ�ɫ�Ͱ�ɫ���Ļ��ơ������������˵���Ҫ����֮һ�����ǽ���“���߾�”��“���߾�”��ҩ�Ƶ�ԭ��֮һ�������߳ƽ�Ǯ���ߣ�Ϊ��������ҩ�ġ��ֲ����ҹ��������ϸ�ʡ���������н��ߣ�Bungarus fasciatus��������к�ɫ�ͻ�ɫ���Ļ��ơ�
�������壩���߿ƣ�Hydrophiidae������ȫ�����������ں����е�ǰ������ߡ����β��⣻�ǿ�λ���DZ�������������ıǰꣻ����խ����ʧ����̥����������Ϊʳ�����ƹ�49�֣��ֲ���ӡ�����̫ƽ����ˮ�����ҹ��غ���16�֣��ֲ�����dz��Ǻ��ߣ�Pelamis platurus�������屳�ڸ��ƣ���Ȼ�ֽ磬�ʱ����Ƹ����ߡ�β������5��10��ڰߣ�β�˺�ɫ�����ܣ���С��ͼ��ද��ֲ����ҹ��Ļƺ����������Ϻ��غ���
������������ƣ�Viperidae�����Ƕ̶��ߣ������ų��������Ĺ��������ɸ���������ͷ�ǵĻ�е����տ�ʱ������ͬ�������ڿ�ǻ����������ʱ���Ǻ�����һ������������ȫ�Ƕ��ߣ��߶�ΪѪѭ���࣬��Ҫ��������Ѫ��ϵͳ��ѪҺ������ͷ�������ѺͶ��۵Ĵ�С���ɷ�Ϊ���ǿƣ�Crotalinae�������ǿƣ�Viperinae���Ͱ�ͷ���ǿƣ�Azemiopinae�������ƹ���180���֣��������ֲ����棬���ǿƷֲ������ޡ�����ŷ�ޣ��������ǿ����࣬�����ǿƵķֲ������ڷ��ޣ�ŷ��������Ҳ�У�����ȱ�����ǿ����ࡣ��ͷ���ǿƽ�1��1�֣�����ͷ��Azemiops feae�����������ҹ���Խ�ϡ��ҹ���������������У�
�������ߣ�Agkistrodon halys��ͷ�Գ������Σ��������ԡ��м��ѡ����۾��⡣������ɫ�仯�ڻҺ�ɫ������ɫ֮�䣬���������źں�ɫԲ��״��ߡ��ۺ��к�ɫ���ߣ����Ϸ����лư�ɫϸ�ơ���Ҫ�Լ�����Ϊʳ����̥����ÿ�β�����2��15�����ֲ������鼰ȫ����
�����������岽�ߣ���Deinagkistrodon acutus���峤��1.5m���Ƕ˼�������Ϸ���������ޭ�ߣ��������ԡ�����Һ�ɫ�����лҰ�ɫ�����η��ߣ������ɫ���кڰߣ��׳ư��ߺ�����ߡ���������Ҫ�ֲ��ڳ������ϵ�����
��������ͷ��Trimeresurus jerdonii��ȫ��1m���ҡ������������������ɫ�����лơ��켰��ɫ�ߵ㣬����ڼ��������γ�һ�нϴ�İ߿顣���ں���1700m��3100m��ɽ��ʯ�ѻ��Ӳ��У��Ǵ��������ľ綾�ߣ������С�����ࡣ��̥��������������Ҷ�ࣨTrimeresurus stejnegeri����ͷ�������Σ���ϸ��������ɫ������Ծ���β��һ�������Ļ�����ߣ����ڱ�ʶ����������˵ľ綾��֮һ�����߶��ֲ��ڻƺ����ϵ�����
������ԭ��Vipera ursini�����ѡ�ͷ��������Ƭ�⻬�������Һ�ɫ������������һ�кں�ɫ���״���ƣ�����а���ɫ�ߵ��ɵ�����2��3�С������ڲ�ԭ�����ֺ�«έ�ԣ�Ҳ���ں���3000m��ɽ�����Իȳ棨ռʳ��������90�����ϣ������档��̥������1��6�����ߣ����ɲ�17�����ֲ����½����ǵ��ص���Ҫ���ߡ�
��
�塢��Ŀ��Crocodiliformes��
��
������Ŀ������2����ȫ���������ң�ͷ���з���Ĵ���������в����ݣ��ߺ������˫���͡�������������ĸ�Ĥ�ߡ�
�����ִ���Ŀ���ﹲ22�֣��ֱ����������ƣ�Crocodilidae�������ƣ�Alligatoridae����ʳ�����ƣ�Ga-vialidae�������Ǵ�������ڷ��ޡ������ޡ������ϲ����ȴ�������ů�������ҹ������С����ε��������������ܺ����������ȥ�ڱ����´��ֲ���Ψһ���š�
������������Alligator sinensis����С������֮һ��ȫ��Լ2m���й��Ž��г�Ϊ�����Ƕ̶��ۣ������ijݴ���ʱǶ�������İ����ڡ�Ƥ�����ʣ����н��ʷ��δ��ۡ�ǰ֫5ָ����֫4ֺ���ڲ�3ָ��ֺ��צ��йֳǻ�����ѡ����ڽ������ߵ�̲�أ�«έ�����ִ����������۷ֱ���Ѩ���ӣ�����ͨ������������ɳ�����10m���ʸ��ӵ��Թ�״��11��������4��Ϊ�����ڣ�4��5�����ѳ�����6��7�½��н�����������ѣ�����Լ20ö�����ʱ�ɴ�52ö�������л���ϰ�ԣ��ѵķ�����Ϊ60��70�죬ҹ����ʳ��ʳ���е���Ҫ�ɷ������ݺ����ϣ�ռ����ʳ����63����Ҳ���㡢�ܡ�Ϻ������С�ȡ�
��
�������������ҹ����ز���Ŀǰ���ķֲ��������ڰ���ʡ�������ϡ�����ɽϵ�Ա�������ش����Լ�����ʡ�������Ͻ�����㽭ʡһ�ǣ�ȫ��Ұ�����岻��500β��Ϊ�ҹ�һ���ص㱣�����1982�꣬�ҹ��ڰ���ʡ�ϲ������dz�������������ֳ�о����ģ������˹��������������������Ѵﵽ��ǧβ֮�࣬Ϊ���ȹ�����ʹ�����Σ������Ѿ�������ˣ������˾��ס�
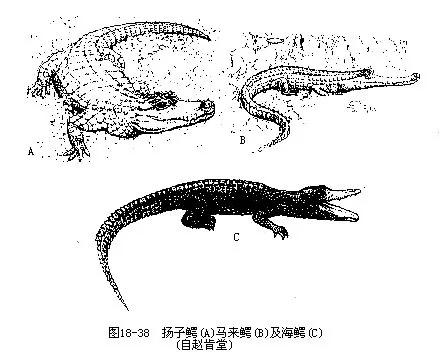
������������Tomistoma schlegelii���Ƿֲ������������ӡ�������ǵĴ��͵�ˮ���ࡣ����һǧ����ǰ���㷺���������ҹ��㶫��������ʡ���������������ڶࡢ������Ϯʳ������ǵ��ص�һ�����ƴ��㶫���ݣ�������ʷ������д���������������ġ���������ָ�ļ�������������������仯���˿��ܼ���������ũ���ƻ���Ϣ����������£�������ǵ�����ɱ¾��������Ԫ���������ڼ���������������Ϊ�ҹ���������Crocodilus porosus�����ҹŽ���Ҳ���ں��ϵ�����ɳȺ��һ��������ˮ���ļ��أ������Ϻ��غ���ȥ�й�����������Ҳ���п��ܵģ�ͼ18—38����
��
������ ���ж���������Ĺ�ϵ
��
һ�����ж�����洦
��
������һ����̬ϵͳ�е��������ж���Ϊ���¶����Ҫ��������̫���ķ�������ά�ֺ�������¡������³´�л�ʵͣ�����Ȼ������Ϊ������Դ��Ӫ����������Ҳ�٣�����������Ĵ���������ͨ��ͬ�����ö�ת��Ϊ���������������侻�������ɴﵽ30����90����ԶԶ�������¶��
������������ж��ﶼ����ʳ����ʳ�࣬���������ͨ��������ʳ���漰���������������������ũ��ҵ����������̬ϵͳ�г䵱�Ŵμ������ߵĽ�ɫ���ݶ�����ϵ���ĵ��飬��βʯ����Ϊ53.69����ʯ����Ϊ51.6�����i��Ϊ24.01�������ֲ�����IJ�ԭɳ����ܵ����ᣬ�ֱ�ﵽ82.14����81.10�����������ж�������ʳ�������ݵ�ʳ�P��������Դ֮һ������̬ϵͳ��������ת�����У��ִ��ڴμ��������ĵ�λ����ˣ����ж����ά��½����̬ϵͳ���ȶ��ԣ��Լ�Ϊ��Ȼ���ṩ����������˵�����в��ɺ��ӵ����á�
����������ʳ������ζ��������Ӫ������֬��22.1����������18�������ж��ְ�����ɷ֣������Ƕ��������̲����β����õ�ʳƷ������������۾��ߡ����ߺͻ�����Ϊԭ���������������߲ˡ����߾ơ����ߵ������������ǵ�������ʳ�������Ե��ȹ�ߣ��������������˿ڵļ��ȣ���Ϊ������̲�ʳƷ�������⡢�군�����š�����������Ƭ���Ļ�֬������̫ƽ�������ൺ������ϲ����ʳ��Ŀǰ�����Ҳ����ԣ��ѵ�����Դ�����ƻ���������Կ��ơ�
������������������Ƥ��Ƥ���ᱡ���������ԣ��������ۣ�������������Ƥ�Ƥ����ƤЬ�������Ǯ���ȹ���Ʒ��Ҳ���������١��ֹġ����ҵ���Ĥ����Ƥ������������
�������ģ�ҽҩ���ҹ�������ҩ������������10���֣������ʢ�����Dz��ڹ����Ĵ�ڻ���ҩ��Ϊ���ݡ����ݸ�Ŀ�������в��������澫Ѫ������ֹ�ȡ��Ʒ�Ӹ���ʡ��������Ĺ��ܡ���������Ϊ������Ʒ�������غ��½������ᣬ���ɹ��л�Į���ᣬ����ʡ������ɳ���ȡ����塢���⡢�ߵ������ɡ��߶�������ҩ���ߵ��ɼӹ����ߵ�����Һ���ߵ���Ƥĩ���ߵ�����Һ���г�ҩ���η�ʪ�ؽ�ʹ�����Զ�̵�Ȳ������ɵ���ҩ�������£���ҩ��ɱ�����Ĺ��ܣ������ƺ������ס���Ѣ���Ѳ���
�����߶��о��������ѧ�м��п���ǰ;��һ����֧���ҹ����߶��о������߶�Ѫ������ƣ������뵽�й��߶������������ۺ����÷��档Ŀǰ���Ƴɵ��۾��߶�ע������б���ȸ���Ч���־õ���ʹ���ã����ڼ�������ת�ư�ʹ��������������ʹ����ʪ�Թؽ�ʹ��������Σ��״����Ȳ��˵ľ�ʹ���������Ե�Ч�������߶�ø���ư�֢Ҳ�յ���һ����Ч�������߶��н�ǿ����Ѫ�ԣ����ڻ���ȱ����Ѫ��Ѫ�Ѳ����ߣ������߶��������Ѫ�Լ����ľֲ�ֹѪ���ҹ�ѧ�ߴ������߶�����ȡ�Ŀ�˨ø�����ٴ���������Ѫ˨��Ѫ˨�����������ס����IJ������ơ�
�������壩�������� ����Եؿ��ڲ��ľ����������������淢������������б�˶��ȣ����к�ǿ�������ԣ���������ڵ���ǰ���ֳ���������Ϊ�����ŷ���ѧ�ķ�չ����ѧ�һ����ݶ����ѵĹ��켰����ص��Ȳ�λ�����ã����о��ɹ�Ӧ�õ������߲�λ���ϣ����Ƴɾ��и߶Ⱦ�ȷ�Ժ����ٷɻ���DZͧ����������β�ߵ���������Ե�װ�õȡ��Ӻ������·�ߵĵ������ƿ��������ƺ����������о���Ҳ������Ϊ����������Žṹ�ı��ǽṹ���ۣ��ڴ��ͽ���������н��֮����
�����ҹ������ж�����Դ�ḻ�������������������������������ز����࣬���ڿ�ѧ�о������Ļ�����������Ҫ���塣
��
�������ߵ�Σ�������˷���
��
������һ�����������ߵ��������ִ�������У�Լ�н�600��Ϊ���ߣ��ҹ�������50�����ҡ�
�����߶���һ�ָ��ӵĵ����ʣ������˺Ͷ������ں������ܰͼ�ѪҺ��ɢ�������ж�֢״��ǰ����������߶����˵�Σ���ϴֱ����۾�������α-���߶����ͺ�����������neurotoxic������������������ʧȥ���ã���ɻ��������ͷ֮��ij嶯�������裬��ʱ���ڵ���������ϵͳ��Զ�������������߶��к���Ѫѭ����hemotozic�����������˿ھ�ʹ��ˮ�ס�����Ƥ�³����ϰߣ����������˥�����������߶�Һ����Ҫ�ɷּ���18—1��
 ��
��
����ͨ���߶��Ķ���ǿ��������߶������ʣ��Լ�����ҧ��ʱ���Ŷ����йء����磬�۾������Ŷ����࣬����ǿ����Ҷ���Ŷ����٣�����Ҳǿ���������Ŷ����٣�������ǿ���˽���Щ������Ч��Ԥ��������ҧ�˺ʹ������ˣ�������Ҫ�Ļ������塣
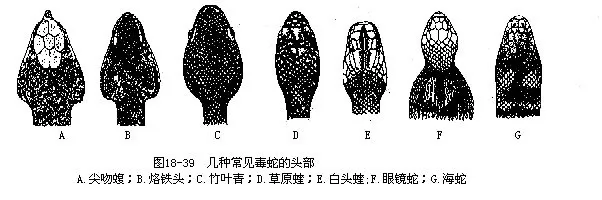
������������������Ҫ���������ڶ����ж��ٺͶ����������������������⣬�������и���ɹ�ʶ�綾�ߺ����ߣ���18—2��ͼ18—39��ͼ18—40����

 ��
��
�����ҹ��Ķ����У���10���������߿Ƶĺ����࣬�����������Σ�������߿ƵĶ���Լ��16�֣����������ں����У�ֻ���غ�������ż����ҧ�IJ���������20�ֶ����У���������13��ʮ�ֺ�����ֲ���������խ�����Զ��Խ�ǿ���ֲ��Ϲ㡢�����϶�;����������˵Ķ��ߣ�ͨ��ֻռ���������1/5���ҡ������Ա��������ڵ���Ҫ���������ߡ������Ĵ����ƹ��ԭ�����ɽ�������ϵ�������Ҫ���������ߡ�������ͼ�������ͷ���ɳ����С������ذ������ϵ�������ɽ���Ļ��е���������ش������ߡ��۾��������ߣ���ɽ�����м��������Ҷ��ȡ�����ɽ���������ϵĻ��ϵ��������ҹ����������������Ҫ�������۾��ߡ������ߡ����ߡ��۾����ߡ�����������ͷ����Ҷ��ȡ���ظ�ԭ����Ҫ�������۾����ߡ���ԭ��ī����Ҷ�ࡢ������Ҷ�༰�˻�����ͷ�ȡ�
�������������˼������ ����ҧ�������صĵ��������������ǺͶ����Ǹ������ݱ�����������ÿ��Լ����ʮ���˱�����ҧ�ˡ�ӡ����1968��������������ߴ�30��40���ˣ������ľ���3����ˡ�ͬ���������Ǵδ�½���ҹ�Ҳ���൱���������˲��ˣ��ɼ�����ҧ������Щ������Σ���Ƿdz����صġ����⣬��ԭ�������ر����紺���ڣ��Ķ��߳�����Ⱥ����˺��������ҹ��½�������������ߡ���ԭ��Ͱ���̩�����ļ�����ȶ��ߣ���Σ������ȫ����ȥ���ߺ���������ƥ��Լռ����������20���������൱�ɹۡ��������ҹ�����ѧ�������뵱��Ⱥ�����ϣ�������̬ѧ��ҩѬ���������ߺ���ȡ�������ɼ���

��
����һ����˵�����ߵ��ж����Ƚϳٻ���������������ҧ�ˣ�ֻ�е������ڿ���ݡ�����ҹ���У�����ȵ���Ӵ�����ʱ�Żᷢ��ҧ���¹ʡ���ˣ����˵IJ�λͨ��������֫�Ľ������£��������֫��ͷ���ز�������������������18�桫30�棬�������ҹ��������ϵ�����7��9�������˷�������ߵļ��ڣ��������ļ�������������է���������������߶�����ѹ�Ͷ�ʪ�ȴ��߾����������ҧ�����ˡ�
�������������ҧ�ˣ�������������Ӧ���������У�ͬʱ���ߴ�����ҽ����Ը��ݶ��ߵ���������ȡ��֢�����Ǽ�Ϊ��Ҫ�ġ�����ȷϵ������ҧ���ͻ����˴�����2�����������ۣ�������˿�������ʹ���ڼ�������������������������Ѹ����չ���ͷ�Χ��ͬʱ���ᷢ��ͷ�Ρ��ۻ����鴤����˯��֢״��
��������ҧ�˵Ľ����ֲ�����ԭ���Ǿ����ų���Һ���ӻ��߶�����ɢ���Լ����ж�֢״��һ��Ӧ�������˿��Ϸ�2cm��10cm���ò�������������ܰͺ;���Ѫ�Ļ�������ÿ��15��20���ӷ��ɲ���1��2���ӣ�����ѪҺѭ�����裬��ɾֲ���֯��������ע�俹�߶�Ѫ��ɽ��������������Ӧ����ˮ����ˮ��0.5��Ũ�ȵĸ��������Һ������ϴ�˿ڡ����⣬����ʹ�������Ŷ����������������ҧ�˲��˲��ô˷������λ����������ų��߶�������������Ҫ��ʱ�ͽ���ҽ���ơ�
�����ҹ����߶����������˷��η�����о�����ȡ���ش�ɾ͡�Ŀǰ�������õ��ۼ���ۿ��߶�Ѫ���α-�ӵ���ø����Чҩ�����������⣬�����ö��ֲ�ҩ�����Ƴɸ�����ҩ�����������˶���ҧ�˵������ʡ�
��
��ϰ��
��
����1.������Ĥ�ѵ���Ҫ���������ڶ����ݻ�ʷ�ϵ����塣
����2.������������Ӧ��½������Ҫ������
����3.�����ִ�������4��Ŀ����Ҫ�Ƶ�����
����4.�ܽ����ж���������Ĺ�ϵ��
����5.���������ߵ������Լ����߷���ԭ��